aún se sabe poco acerca de los mecanismos que regulan el proceso de la cicatrización, al grado de que con los recursos actuales, el cirujano se limita a no interferir y, en el mejor de los casos, a favorecer el proceso biológico que hace sanar las heridas sin que se conozca hasta el momento cómo regular su evolución.
Para el cirujano, la herida que atiende llega a sanar en un proceso continuo y predecible, que finaliza cuando la lesión «cicatriza» y significa que se restableció la integridad física. En la investigación básica se considera el mismo proceso como una cascada armónica de hechos fisiológicos, a menudo difíciles de relacionar entre sí, que culmina con la creación de un nuevo órgano, la cicatriz, la cual evoluciona con el tiempo y continuará siendo objeto de estudio.
En realidad no existe ninguna incongruencia en estos dos puntos de vista, puesto que cirujano y paciente observan efectos macroscópicos de un fenómeno biológico, en tanto que el investigador básico estudia los hechos que ocurren a nivel microscópico y molecular
LESIONES son los daños que llagan a sufrir los tejidos por algún tipo de agresión ya sea física, química o biológica. presentan alteraciones más o menos profundas de la forma y función de los tejidos y órganos, que varían según el tipo de agente agresor, la magnitud del daño y la naturaleza del tejido o de los tejidos que resultaron afectados
HERIDAS son lesiones ocasionadas por traumatismo mecánico en las que se observa rotura o interrupción de la continuidad de los tejidos blandos, y cuando el tejido lesionado es rígido o semirrígido, a la solución de la continuidad se le conoce como fractura
Los puntos clave de la «cicatrización» :
l. El proceso de sanar en los tejidos lesionados es una compleja cascada de eventos celulares, los cuales son coordinados por mediadores solubles y conducen a su restitución física y funcional.
II. los tejidos tiene características propias del proceso, pero todos los tejidos sanas por mecanismos similares cursando por las fases de: inflamación, migración celular, proliferación, deposito de matriz y remodelación.
III. El cirujano debe conocer los factores locales, los factores sistémicos y las causas técnicas que pueden obstaculizar la evolución normal del proceso
IV. Los resultados óptimos se obtienen con la evaluación integral del paciente, de la herida y la aplicación de las mejores técnicas de práctica clínica.
V. La cicatrización anormal por exceso o por defecto plantea problemas clínicos en los que la genética, los factores del paciente y una buena técnica son determinantes.
Clasificación de las heridas según su profundidad
l. Excoriacion. Lesión superficial que afecta la epidermis y en general cicatriza regenerando en forma íntegra el epitelio, sin dejar huella visible.
2. Herida superficial. Es aquella que involucra la piel y al tejido adiposo hasta la aponeurosis.
3. Herida profunda. Afecta los planos superficiales, la aponeurosis, el músculo y puede lesionar vasos, nervios y tendones.
4. Herida penetrante. Herida que lesiona los planos superficiales y llega al interior de las grandes cavidades. se les llama penetrante al abdomen, penetrante al tórax y penetrante al cráneo. En ocasiones hay dobles penetrantes, por ejemplo, al tórax y al abdomen.
Fases de la cicatrización
Hemostasia y fase inflamatoria
Esta respuesta es considerada como la preparación de un sustrato o base orgánica y tisular que tiene como fin la curación y presupone una defensa contra otras lesiones o invasiones futuras, así como también la liberación de factores solubles quimiotácticos que controlan la permeabilidad de los vasos y otros que atraen o atrapan células. En esta fase domina el flujo de elementos hemáticos al sitio de la lesión con la liberación de citocinas y otros mediadores de la inflamación. A continuación se listan los elementos que participan en esta fase
plaquetas: Las plaquetas contienen al menos tres tipos de organelos: gránulos, cuerpos densos y lisosomas. Los gránulos contienen factores de crecimiento, así como factor transformador del crecimiento y fibronectina. Los cuerpos densos almacenan aminas vasoactivas (serotonina) y los lisosomas contienen proteasas. Durante el proceso de coagulación las plaquetas liberan fibrinógeno, fibronectinas, trombospondina y factor de Von Willebrand. Todos estos elementos intervienen en la adhesión de las plaquetas a la colágena que quedó expuesta en la herida.
coagulacion: El coágulo de fibrina no sólo tiene la función de hacer hemostasia, sino que junto con la fibronectina forma el armazón sobre el que migrarán los monocitos, fibroblastos y queratinocitos
Leucocitos: Con el estímulo de los productos de la cascada de la coagulación, los neutrófilos son las primeras células nucleadas en llegar. Esta migración es resultado de un proceso complejo que facilitan el paso de los neutrófilos a través de las células endoteliales de los capilares por medio de un mecanismo conocido como diapédesis Al principio, monocitos y macrófagos son atraídos por los mismos mediadores químicos que estimularon a los neutrófilos, después por quimiotácticos específicos y en poco tiempo se convierten en las células dominantes del proceso inflamatorio
histamina, serotonina, sistema de cininas y proteínas séricas sustancias producen estímulos que modifican la actividad y la permeabilidad vascular en el lado venoso de los capilares
Fase proliferativa
epitelización: La respuesta de las células epidérmicas inicia dentro de las primeras 24 horas de sufrida la lesión. A las 12 horas de perder contacto con sus homólogos vecinos, los queratinocitos de los bordes de la herida y de los folículos pilosos o de las glándulas sebáceas se aplanan, forman filamentos de actina en su citoplasma, emiten prolongaciones semejantes a pseudópodos y emigran La mitosis tarda más en iniciar que la migración, por lo general de 48 a 72 horas después de la lesión. Las células se multiplican y movilizan hasta que entran en contacto con otras células epiteliales, momento en el cual se inhiben. Entonces las células toman una apariencia similar a la que tuvieron en condiciones basales y, a medida que maduran, forman queratina un vez terminada la migración se sintetiza la membrana basal a través de adhesión con el colágeno.
Angiogénesis: Los monocitos y los macrófagos producen factores que inducen la formación de nuevos vasos por los que se transporta oxígeno y nutrientes a la herida, y secretan sustancias biológicamente activa. Las células que intervienen en el proceso de angiogénesis son las endoteliales. Los nuevos vasos se originan como capilares que brotan a los lados de los pequeños vasos a manera de respuesta a los factores angiógenos; emiten pseudópodos a través de la lámina basal y se proyectan al espacio perivascular. Después se dividen, se forman vacuolas y se fusionan para crear un nuevo lumen
Matriz de la herida (sustancia fundamental): El tejido herido no sólo está formado por células; también es preciso considerar el espacio extracelular, el cual está lleno de macromoléculas que forman una matriz compuesta por proteínas fibrosas embebidas en gel de polisacáridos, hidratados y secretados por los fibroblastos.
fibroplasia y síntesis de colágeno: Los fibroblastos aparecen entre 48 y 72 horas después de ocasionada la herida, y son estimulados y regulados por factores quimiotácticos complejos que provienen de los macrófagos presentes en la herida Son los protagonistas de la proliferación y, junto con los nuevos vasos y la matriz, forman el tejido llamado de «granulación»
contraccion de la herida: Es una disminución gradual del área de la herida por retracción de la masa central del tejido de granulación. Las fuerzas contráctiles producidas por este tejido son resultado de la acción de los miofibroblastos que contienen proteínas contráctiles El fenómeno de contracción en las heridas profundas es fundamental y se le debe 40% de la disminución del tamaño de la lesión.
Fase de Remodelación
Cuando ha sido reparada la rotura de la continuidad de los tejidos, el estímulo angiógeno disminuye en intensidad y, al parecer, como respuesta a las tensiones elevadas de oxígeno en los tejidos se inicia un periodo en el que la herida madura, la cual presenta remodelación morfológica, también disminuyen la hiperemia y su vascularidad, asimismo se reorganiza el tejido fibroso neoformado
Hormonas corticosuprarrenales Las dos glándulas suprarrenales, con un peso aproximado de 4 g cada una, se hallan en los polos superiores de los riñones. cada glándula se compone de dos porciones diferentes, la médula suprarrenal y la corteza suprarrena La médula suprarrenal, que ocupa el 20% central de la glándula, se relaciona desde el punto de vista funcional con el sistema nervioso simpático; secreta las hormonas adrenalina y noradrenalina en respuesta a la estimulación simpática La corteza suprarrenal secreta un grupo completamente diferente de hormonas, llamadas corticoesteroides. Todas estas hormonas se sintetizan a partir del esteroide colesterol y todas poseen una fórmula química parecida.
Corticoesteroides: mineralocorticoides, glucocorticoides y andrógenos
Los mineralocorticoides reciben este nombre porque afectan sobre todo a los electrólitos (los «minerales») del compartimiento extracelular, especialmente al sodio y al potasio. Los glucocorticoidesse denominan así porque poseen efectos importantes de aumento de la glucemia. Además, influyen en el metabolismo de las proteínas y de los lípidos, con efectos tan importantes para la función del organismo como los que producen sobre el metabolismo de los hidratos de carbono. Además de estas hormonas, produce pequeñas cantidades de hormonas sexuales, en particular de andrógenos, que inducen los mismos efectos que la hormona sexual masculina testosterona
Síntesis y secreción de hormonas corticosuprarrenales La corteza suprarrenal tiene tres capas diferentes
La zona glomerular, una capa delgada de células situada inmediatamente por debajo de la cápsula, contribuye con casi el 15% a la corteza suprarrenal. Estas células son las únicas de la glándula suprarrenal capaces de secretar cantidades importantes de aldosterona porque contienen la enzima aldosterona sintetasa, necesaria para la síntesis de la hormona
La zona fascicular, la zona media y más ancha, representa casi el 75% de la corteza suprarrenal secreta los glucocorticoides cortisol y corticosterona
La zona reticular, la capa más profunda de la corteza, secreta los andrógenos suprarrenales deshidroepiandrosterona (DHEA) y androstenodiona
Funciones de los mineralocorticoides: aldosterona
La deficiencia de mineralocorticoides provoca pérdidas renales intensas de cloruro sódico e hiperpotasemia La pérdida completa de la secreción corticosuprarrenal puede causar la muerte en un plazo de 3 días a 2 semanas, salvo que la persona reciba un tratamiento salino intensivo o la inyección de mineralocorticoides. Sin mineralocorticoides, la concentración del ion potasio del líquido extracelular experimenta un gran ascenso, el sodio y el cloruro desaparecen enseguida del organismo y el volumen total del líquido extracelular y el volumen de sangre se reducen mucho. Pronto se desarrolla un descenso del gasto cardíaco, que evoluciona a un estado de shock, seguido de la muerte
Efectos renales y circulatorios de la aldosterona La aldosterona aumenta la reabsorción tubular renal del sodio y la secreción de potasio
la aldosterona conserva el sodio en el líquido extracelular y aumenta la eliminación urinaria de potasio. El incremento de la concentración de aldosterona del plasma puede reducir de forma pasajera la pérdida urinaria de sodio y dejarla en unos pocos miliequivalentes al día. Al mismo tiempo, las pérdidas de potasio por la orina se multiplican transitoriamente. Así pues, el efecto neto del exceso de aldosterona en el plasma consiste en un aumento de la cantidad total de sodio en el líquido extracelular y un descenso de la de potasio
El exceso de aldosterona aumenta la secreción tubular de iones hidrógeno, con la consiguiente alcalosis leve La aldosterona no solo induce la secreción tubular de potasio, que se intercambia por el sodio reabsorbido en las células principales de los tubos colectores renales, sino que también provoca una secreción de iones hidrógeno, intercambiados por potasio, por parte de las células intercaladas de los tubos colectores corticales, como se expone en los capítulos 28 y 31. Como es lógico, la concentración de iones hidrógeno disminuye en el líquido extracelular. Este efecto suele provocar una alcalosis metabólica
Regulación de la secreción de aldosterona La regulación de la secreción de aldosterona está tan íntimamente ligada al control de las concentraciones de electrólitos en el líquido extracelular, el volumen del líquido extracelular, el volumen sanguíneo, la presión arterial y muchos aspectos especiales
Se conocen los siguientes cuatro factores que desempeñan una función esencial para la regulación de la aldosterona:
El incremento de la concentración de iones potasio en el líquido extracelular aumenta mucho la secreción de aldosterona.
El aumento de la concentración de angiotensina II en el líquido extracelular también incrementa mucho la secreción de aldosterona.
El incremento de la concentración de iones sodio en el líquido extracelular apenas reduce la secreción de aldosterona.
Se necesita ACTH de la adenohipófisis para que haya secreción de aldosterona, aunque su efecto regulador sobre la velocidad de secreción es mínimo en la mayoría de los trastornos fisiológicos.
Funciones de los Glucocorticoides
Al menos el 95% de la actividad glucocorticoide de las secreciones corticosuprarrenales se debe a la secreción de cortisol, también conocido como hidrocortisona. Por último, la corticosterona posee una actividad glucocorticoide pequeña, pero importante
Efectos del cortisol sobre el metabolismo de los hidratos de carbono Estimulación de la gluconeogenia El efecto metabólico más conocido del cortisol y de otros glucocorticoides consiste en estimular la gluconeogenia (es decir, la formación de hidratos de carbono a partir de las proteínas y de otras sustancias) en el hígado; el ritmo de gluconeogenia se eleva, a menudo, entre 6 y 10 veces. pues la hidrocortisona estimula la formacion de enzimas que participan en la conversion de aminoacidos en glucosa. ademas de movilizar los aminoacidos de tejidos extrahepaticos, especialmente de los musculos. importante el antagonismo que tiene con la insulina.
Disminución de la utilización celular de la glucosa El cortisol también reduce, aunque en grado moderado, la utilización de glucosa por la mayoría de las células del cuerpo. Aunque se desconoce la causa exacta de este descenso, un efecto importante del cortisol es la reducción de la translocación de los transportadores de glucosa GLUT-4 en la membrana celular, en especial en las células del músculo esquelético, lo que conduce a resistencia a la insulina
Efectos del cortisol sobre el metabolismo de las proteínas Reducción de las proteínas celulares Uno de los principales efectos del cortisol sobre los sistemas metabólicos del organismo consiste en el descenso de los depósitos de proteínas de la práctica totalidad de las células del organismo, con excepción de las del hígado. Esta reducción se debe tanto al descenso de la síntesis como a un mayor catabolismo de las proteínas ya existentes dentro de las células.
lo contrario sucede en el higado en donde la hidrocortisona estimula un ascenso en la formacion de proteinas hepaticas al transportar mas aminoacidos, de esta forma se libera mas proteinas al plasma sanguineo, pero se reduce en las demás células del organismo.
Efectos del cortisol sobre el metabolismo de las grasas Movilización de los ácidos grasos De forma muy similar a la movilización de aminoácidos del músculo, el cortisol moviliza a los ácidos grasos del tejido adiposo. Esta movilización aumenta la concentración de ácidos grasos libres en el plasma, lo que aumenta también la utilización de los ácidos grasos con fines energéticos. Al parecer, el cortisol ejerce asimismo un efecto directo que potencia la oxidación de los ácidos grasos en el interior de la célula.
Efectos antiinflamatorios de las concentraciones altas de cortisol Cuando un tejido sufre daños a causa de un traumatismo, una infección bacteriana o cualquier otra causa, suele «inflamarse».
Síntesis y secreción de las hormonas metabólicas tiroideas
Alrededor del 93% de las hormonas con actividad metabólica secretadas por la glándula tiroides corresponde a tiroxina y el 7% restante, a triyodotironina. No obstante, con el tiempo, casi toda la tiroxina se convierte en triyodotironina en los tejidos, por lo que ambas desempeñan funciones importantes. Estas funciones son cualitativamente similares, aunque difieren en la rapidez y la intensidad de la acción. La triyodotironina es unas cuatro veces más potente que la tiroxina, si bien se detecta una cantidad mucho menor en la sangre y su duración es más breve.
Anatomía fisiológica de la glándula tiroides la glándula tiroides se compone de un elevado número de folículos cerrados (100 a 300 μm de diámetro), que están repletos de una sustancia secretora denominada coloide y revestidos por células epiteliales cúbicas que secretan a la luz de los folículos. El componente principal del coloide es una glucoproteína de gran tamaño, la tiroglobulina, cuya molécula contiene las hormonas tiroideas. Cuando la secreción se encuentra en los folículos, la sangre debe absorberla de nuevo a través del epitelio folicular para que pueda actuar en el organismo. El flujo sanguíneo por minuto de la glándula tiroides equivale a unas cinco veces su peso, lo que supone un aporte sanguíneo comparable al de cualquier otra región del organismo, con la posible excepción de la corteza suprarrenal.
El yoduro es necesario para la formación de tiroxina Para formar una cantidad normal de tiroxina se precisan al año unos 50 mg de yodo (ingerido en forma de yoduros) o el equivalente a 1 mg/semana. Para impedir la deficiencia de yodo, se añade una parte de yoduro sódico por cada 100.000 partes de cloruro sódico a la sal de mesa común.
Bomba de yoduro: el simportador del yoduro de sodio (atrapamiento de yoduro) La primera etapa de la formación de las hormonas tiroideas, consiste en el transporte de los yoduros desde la sangre hasta las células y los folículos de la glándula tiroides. La membrana basal de estas células posee la capacidad específica de bombear de forma activa el yoduro al interior celular. Este bombeo se consigue mediante la acción de un simportador del yoduro de sodio, que cotransporta el ion yoduro a lo largo de dos iones sodio a través de la membrana basolateral (plasma) a la célula
Oxidación del ion yoduro El primer paso crítico para la formación de las hormonas tiroideas consiste en la conversión de los iones yoduro en una forma oxidada del yodo, bien en yodo naciente (I ), bien en I3, que luego puede combinarse directamente con el aminoácido tirosina. La oxidación del yodo depende de la enzima peroxidasa y su peróxido de hidrógeno acompañante, que constituyen un potente sistema capaz de oxidar los yoduros
Almacenamiento de la tiroglobulina La glándula tiroides es la única glándula endocrina que posee la capacidad de almacenar grandes cantidades de hormona. Una vez finalizada la síntesis de las hormonas tiroideas, cada molécula de tiroglobulina contiene hasta 30 moléculas de tiroxina y algunas de triyodotironina. De esta forma, los folículos pueden almacenar una cantidad de hormona tiroidea suficiente para cubrir las necesidades normales del organismo durante 2 o 3 meses. Por consiguiente, cuando cesa la síntesis de hormona tiroidea, los efectos fisiológicos de la deficiencia tardan varios meses en aparecer.
Secreción diaria de tiroxina y de triyodotironina En condiciones normales, alrededor del 93% de la hormona tiroidea liberada por la glándula tiroides corresponde a tiroxina y solo el 7% es triyodotironina. No obstante, en los días siguientes, la mitad de la tiroxina se desyoda con lentitud y forma más triyodotironina. Por consiguiente, la hormona liberada en última instancia a los tejidos y empleada por ellos es sobre todo la triyodotironina y, en concreto, se generan unos 35 μg de triyodotironina diarios
Transporte de tiroxina y triyodotironina a los tejidos La tiroxina y la triyodotironina están unidas a proteínas plasmáticas Cuando acceden a la sangre, más del 99% de la tiroxina y la triyodotironina se combina de inmediato con diversas proteínas plasmáticas, todas ellas sintetizadas por el hígado. Estas proteínas son, ante todo, la globulina fijadora de la tiroxina y, en menor medida, la prealbúmina y la albúmina fijadora de la tiroxina.
Comienzo lento y acción prolongada de las hormonas tiroideas Cuando se inyecta una cantidad elevada de tiroxina a una persona, no se percibe ningún efecto sobre el metabolismo durante 2 o 3 días, lo que demuestra la existencia de un período prolongado de latencia que precede a la actividad de la tiroxina. Cuando esta actividad comienza, luego aumenta de forma progresiva y alcanza su máximo valor en 10-12 días, para descender después, con una semivida de unos 15 días. Parte de la actividad persiste entre 6 semanas y 2 meses.
Funciones fisiológicas de las hormonas tiroideas
Las hormonas tiroideas aumentan la transcripción de una gran cantidad de genes
El efecto general de las hormonas tiroideas consiste en la activación de la transcripción nuclear de un gran número de genes. Por consiguiente, en casi todas las células del organismo se sintetiza una elevada proporción de enzimas proteicas, proteínas estructurales, proteínas transportadoras y otras sustancias. El resultado neto es un aumento generalizado de la actividad funcional de todo el organismo.
Las hormonas tiroideas aumentan la actividad metabólica celular Las hormonas tiroideas incrementan las actividades metabólicas de casi todos los tejidos del organismo. El metabolismo basal se incrementa entre el 60 y el 100% por encima de su valor normal cuando las concentraciones hormonales son altas. La velocidad de utilización de los alimentos como fuente de energía se encuentra muy acelerada. Aunque la síntesis de proteínas aumenta, también lo hace el catabolismo proteico. La velocidad de crecimiento de las personas jóvenes experimenta una gran aceleración. Los procesos mentales se estimulan y las actividades de las demás glándulas endocrinas se potencian.
Efecto de las hormonas tiroideas sobre el crecimiento En la especie humana, el efecto de la hormona tiroidea sobre el crecimiento se manifiesta sobre todo en los niños en edad de desarrollo. En los niños hipotiroideos, la velocidad de crecimiento es mucho más lenta, mientras que los hipertiroideos a menudo experimentan un crecimiento esquelético excesivo, por lo que son bastante más altos de lo que les correspondería según su edad
Un efecto importante de la hormona tiroidea consiste en el estímulo del crecimiento y del desarrollo del cerebro durante la vida fetal y en los primeros años de vida posnatal. Si el feto no posee cantidades suficientes de hormona tiroidea, el crecimiento y la maduración del cerebro antes y después del nacimiento se retrasarán y su tamaño será más pequeño de lo normal. Si no se aplica un tratamiento tiroideo específico en los primeros días o semanas de la vida, el niño que carece de glándula tiroides presentará un retraso mental permanente.
La TSH adenohipofisaria incrementa la secreción tiroidea Esta hormona, incrementa la secreción de tiroxina y de triyodotironina por la glándula tiroides. Los efectos que ejerce sobre esta glándula son los siguientes:
Eleva la proteólisis de la tiroglobulina que se encuentra almacenada en los folículos, con lo que se liberan hormonas tiroideas a la sangre circulante y disminuye la sustancia folicular.
Incrementa la actividad de la bomba de yoduro, que favorece el «atrapamiento del yoduro» por las células glandulares, elevando en ocasiones la relación entre las concentraciones intra- y extracelular de yodo en la sustancia glandular hasta ocho veces por encima de los valores normales.
Intensifica la yodación de la tirosina para formar hormonas tiroideas.
Aumenta el tamaño y la actividad secretora de las células tiroideas.
Incrementa el número de células tiroideas y transforma las células cúbicas en cilíndricas e induce el plegamiento del epitelio tiroideo en el interior de los folículos.
Efectos del frío y de otros estímulos nerviosos sobre la secreción de TRH y TSH Uno de los estímulos más conocidos de la secreción de TRH por el hipotálamo y, por consiguiente, de la secreción de TSH por la adenohipófisis es la exposición de un animal al frío. Este efecto obedece casi con toda seguridad a la excitación de los centros hipotalámicos encargados de controlar la temperatura corporal. La exposición de las ratas al frío intenso durante varias semanas incrementa la producción de hormonas tiroideas hasta más del 100% de su valor normal y eleva el metabolismo basal hasta en un 50%.
Adenohipófisis y neurohipófisis La hipófisis, denominada también glándula pituitaria, es una pequeña glándula de alrededor de 1 cm de diámetro y 0,5-1 g de peso, situada en la silla turca (una cavidad ósea de la base del cráneo) y unida al hipotálamo mediante el tallo hipofisario. Desde una perspectiva fisiológica, la hipófisis se divide en dos partes bien diferenciadas: el lóbulo anterior o adenohipófisis y el lóbulo posterior o neurohipófisis. Entre estas dos partes existe una pequeña zona poco vascularizada y denominada parte intermedia, mucho menos desarrollada en la especie humana y mucho más grande y funcional en algunos animales
Desde el punto de vista embriológico, las dos porciones de la hipófisis tienen procedencias diferentes: la adenohipófisis deriva de la bolsa de Rathke, una invaginación embrionaria del epitelio faríngeo, y la neurohipófisis lo hace de una evaginación de tejido nervioso del hipotálamo. El origen de la adenohipófisis en el epitelio faríngeo explica la naturaleza epitelial de sus células, mientras que el origen de la neurohipófisis en el tejido nervioso justifica la presencia de abundantes células de tipo glial en esta glándula.
La adenohipófisis secreta seis hormonas peptídicas necesarias y otras de menor importancia, mientras que la neurohipófisis sintetiza dos hormonas peptídicas importantes. Las hormonas de la adenohipófisis intervienen en el control de las funciones metabólicas de todo el organismo
La adenohipófisis contiene diversos tipos celulares que sintetizan y secretan hormonas De ordinario, existe un tipo celular por cada hormona principal formada en la adenohipófisis. Mediante el uso de tinciones especiales a los anticuerpos de gran afinidad que se unen a cada una de las hormonas
Somatótropas: hormona del crecimiento humana (GH).
Corticótropas: corticotropina (ACTH).
Tirótropas: tirotropina (TSH).
Gonadótropas: hormonas gonadótropas, es decir, la hormona luteinizante (LH) y la hormona foliculoestimulante (FSH).
Lactótropas: prolactina (PRL).
El hipotálamo controla la secreción hipofisaria
La secreción de la neurohipófisis está controlada por las señales nerviosas que se originan en el hipotálamo y terminan en la neurohipófisis. Por el contrario, la secreción de la adenohipófisis está controlada por hormonas llamadas hormonas (o factores) de liberación y de inhibición hipotalámicas; estas se sintetizan en el propio hipotálamo y pasan a la adenohipófisis (como se muestra en la figura a través de minúsculos vasos sanguíneos denominados vasos porta hipotalámico-hipofisarios. Estas hormonas liberadoras e inhibidoras actúan sobre las células glandulares de la adenohipófisis y rigen su secreción
Sistema porta hipotalámico-hipofisario de la adenohipófisis La adenohipófisis es una glándula muy vascularizada que dispone de amplios senos capilares entre las células glandulares. Casi toda la sangre que penetra en estos senos atraviesa en primer lugar otro lecho capilar del hipotálamo inferior. A continuación, la sangre fluye a través de unos diminutos vasos porta hipotalámico-hipofisarios y accede a los senos adenohipofisarios. Unas pequeñas arterias penetran en la eminencia media y otros vasos de pequeño calibre regresan a su superficie, donde se unen formando el sistema porta hipotalámico-hipofisario. Estos vasos descienden a lo largo del tallo hipofisario y riegan los senos adenohipofisarios.
Las hormonas liberadoras e inhibidoras hipotalámicas controlan la secreción de la adenohipófisis La función de las hormonas liberadoras e inhibidoras consiste en controlar la secreción hormonal de la adenohipófisis. En el control de la mayoría de las hormonas adenohipofisarias intervienen sobre todo los factores liberadores, pero en lo que concierne a la prolactina, el mayor control lo ejerce probablemente una hormona hipotalámica inhibidora. Las principales hormonas liberadoras e inhibidoras hipotalámicas, son las siguientes:
Tiroliberina u hormona liberadora de tirotropina (TRH), que induce la liberación de tirotropina.
Corticoliberina u hormona liberadora de corticotropina (CRH), que produce la liberación de corticotropina.
Somatoliberina u hormona liberadora de la hormona del crecimiento (GHRH), que produce la liberación de hormona del crecimiento, y hormona inhibidora de la hormona del crecimiento (GHIH), denominada también somatostatina, que inhibe la liberación de la hormona del crecimiento.
Gonadoliberina u hormona liberadora de las gonadotropinas (GnRH), que produce la liberación de dos hormonas gonadótropas: las hormonas luteinizante (LH) y foliculoestimulante (FSH).
Hormona inhibidora de la prolactina (PIH), que inhibe la secreción de prolactina.
La hormona del crecimiento ejerce varios efectos metabólicos
Además de afectar al crecimiento general, la hormona del crecimiento ejerce múltiples efectos metabólicos específicos: 1) aumenta la síntesis proteica en casi todas las células del organismo; 2) favorece la movilización de los ácidos grasos del tejido adiposo, incrementa la cantidad de ácidos grasos libres en la sangre y potencia el uso de los ácidos grasos como fuente de energía, y 3) disminuye la cantidad de glucosa utilizada en todo el organismo. Así pues, la hormona del crecimiento estimula la formación de proteínas, la utilización de los depósitos de lípidos y la conservación de los hidratos de carbono.
La hormona del crecimiento estimula el crecimiento del cartílago y el hueso
Este ocurre como consecuencia de los múltiples efectos que ejerce la hormona del crecimiento sobre el hueso, entre los que destacan: 1) aumento del depósito de proteínas por acción de las células condrocíticas y osteógenas inductoras del crecimiento óseo; 2) la mayor velocidad de reproducción de estas células, y 3) un efecto específico consistente en la conversión de los condrocitos en células osteógenas, con lo que se produce el depósito específico de hueso nuevo.
La hormona del crecimiento ejerce muchos de sus efectos a través de sustancias intermedias denominadas somatomedinas
la hormona del crecimiento actúa sobre el hígado (y en menor medida sobre otros tejidos) para formar pequeñas proteínas denominadas somatomedinas que, a su vez, ejercen un potente efecto estimulador de todos los aspectos del crecimiento óseo. Muchos de estos efectos de las somatomedinas sobre el crecimiento se asemejan a los de la insulina. Por consiguiente, las somatomedinas reciben también el nombre de factores del crecimiento seudoinsulínicos (IGF). Se han aislado al menos cuatro somatomedinas, pero la más importante de ellas es la somatomedina C (denominada también factor de crecimiento parecido a la insulina I o IGF-I). El peso molecular de la somatomedina C oscila en torno a 7.500 y su concentración plasmática guarda una estrecha correlación con la velocidad de secreción de la hormona del crecimiento.
Regulación de la secreción de hormona del crecimiento Después de la adolescencia, la secreción disminuye lentamente con la edad y, en última instancia, alcanza el 25% del nivel de la adolescencia a una edad muy avanzada. La secreción de hormona del crecimiento sigue un patrón pulsátil, con ascensos y descensos. No se conocen los mecanismos exactos que controlan su secreción, pero existen diversos factores relacionados con la nutrición o el estrés que la estimulan: 1) la inanición, en especial cuando existe un déficit grave de proteínas; 2) la hipoglucemia o baja concentración sanguínea de ácidos grasos; 3) el ejercicio; 4) la excitación; 5) los traumatismos, y 6) la grelina, una hormona secretada por el estómago antes de las comidas. Asciende de forma característica durante las dos primeras horas de sueño profundo,
Anomalías de la secreción de hormona del crecimiento
Insuficiencia panhipofisaria (panhipopituitarismo) El término insuficiencia panhipofisaria o panhipopituitarismo hace referencia a una secreción reducida de todas las hormonas adenohipofisarias. Puede ser congénita (presente desde el nacimiento) o aparecer de forma repentina o progresiva en cualquier momento de la vida, casi siempre debido a un tumor hipofisario que destruye esta glándula.
Enanismo Casi todos los casos de enanismo se deben a una deficiencia generalizada de la secreción de la adenohipófisis (panhipopituitarismo) durante la infancia. En general, todas las partes del organismo se desarrollan de forma proporcionada, pero la velocidad de desarrollo es mucho menor. Los niños de 10 años presentan el desarrollo corporal de uno de 4 o 5 y cuando llegan a los 20 años tienen el desarrollo corporal de un niño de 7 a 10 años.
Gigantismo En ocasiones, las células acidófilas de la glándula adenohipofisaria productoras de hormona del crecimiento se tornan hiperactivas y a veces llegan a originarse tumores acidófilos en la glándula. Como resultado, se sintetizan grandes cantidades de hormona del crecimiento. Todos los tejidos del organismo crecen con rapidez, incluidos los huesos. Si la enfermedad aparece antes de la adolescencia, es decir, antes de que las epífisis de los huesos largos se fusionen a las diáfisis, la persona crecerá hasta convertirse en un gigante de 2,5 m
Acromegalia Cuando el tumor acidófilo aparece después de la adolescencia, es decir, cuando las epífisis de los huesos largos se han soldado ya con las diáfisis y todo crecimiento posterior es imposible, los huesos aumentarán de grosor, al igual que los tejidos blandos
Filtración glomerular: el primer paso para la formación de orina
El primer paso en la formación de orina es la filtración de grandes cantidades de líquidos a través de los capilares glomerulares en la cápsula de Bowman, casi 180 l al día. La mayor parte de este filtrado se reabsorbe, lo que deja únicamente 1 l aproximadamente de líquido para su excreción al día, si bien la tasa de excreción renal de líquidos puede ser muy variable dependiendo de la ingestión de líquidos. La alta tasa de filtración glomerular depende de la alta tasa de flujo sanguíneo renal, así como de las propiedades especiales de las membranas de los capilares glomerulares
Composición del filtrado glomerular Como la mayoría de los capilares, los capilares glomerulares son relativamente impermeables a las proteínas, de manera que el líquido filtrado (llamado filtrado glomerular) carece prácticamente de proteínas y elementos celulares, incluidos los eritrocitos. Las concentraciones de otros constituyentes del filtrado glomerular, como la mayoría de las sales y moléculas orgánicas, son similares a las concentraciones en el plasma. Las excepciones a esta generalización son algunas sustancias con un peso molecular bajo, como el calcio y los ácidos grasos, que no se filtran libremente porque están unidas parcialmente a las proteínas plasmáticas.
Membrana capilar glomerular La membrana capilar glomerular es similar a la de otros capilares, excepto en que tiene tres capas principales (en lugar de las dos habituales): 1) el endotelio del capilar; 2) una membrana basal, y 3) una capa de células epiteliales (podocitos) rodeando a la superficie externa de la membrana basal capilar
La capacidad de filtración de los solutos se relaciona inversamente con su tamaño La membrana capilar glomerular es más gruesa que la de la mayoría de los otros capilares, pero es también mucho más porosa y por tanto filtra líquido con mayor intensidad. A pesar de la elevada filtración, la barrera de filtración glomerular filtra de modo selectivo las moléculas basándose en su tamaño y en su carga eléctrica.
Las moléculas grandes con carga negativa se filtran con menor facilidad que las moléculas con el mismo tamaño molecular y cargas positivas El diámetro molecular de la proteína plasmática albúmina es solo de unos 6 nm, mientras que los poros de la membrana glomerular tienen unos 8 nm (80 angstroms). Sin embargo, la albúmina no se filtra por su carga negativa y la repulsión electrostática ejercida por las cargas negativas de los proteoglucanos de la pared capilar glomerular.
Determinantes de la FG
La FG está determinada por: 1) la suma de las fuerzas hidrostática y coloidosmótica a través de la membrana glomerular, que da lugar a la presión de filtración neta, y 2) el coeficiente glomerular Kf En una fórmula matemática, la FG es igual al producto del Kf y de la presión de filtración neta
El aumento del coeficiente de filtración capilar glomerular incrementa la FG El Kf es una medida del producto de la conductividad hidráulica y el área superficial de los capilares glomerulares. Dado que la FG total en los dos riñones es de unos 125 ml/min y la presión de filtración neta 10 mmHg, el Kf normal se calcula en unos 12,5 ml/min/mmHg de presión de filtración
El aumento de la presión hidrostática en la cápsula de Bowman reduce la FG El aumento de la presión hidrostática en la cápsula de Bowman reduce la FG, mientras que reducir la presión aumenta la FG. Pero los cambios en la presión de la cápsula de Bowman no son normalmente un mecanismo importante de regulación de la FG
El aumento de la presión coloidosmótica capilar glomerular reduce la FG A medida que la sangre pasa desde la arteriola aferente a través de los capilares glomerulares hasta las arteriolas eferentes, la concentración plasmática de las proteínas aumenta alrededor de un 20%
Flujo sanguíneo renal y consumo de oxígeno
Con respecto al gramo de peso, los riñones consumen normalmente el doble de oxígeno que el encéfalo, pero tienen casi siete veces más flujo sanguíneo. Luego el oxígeno transportado a los riñones supera con mucho sus necesidades metabólicas, y la extracción arteriovenosa de oxígeno es relativamente baja comparada con la de la mayor parte de los restantes tejidos. Una gran fracción del oxígeno consumido por los riñones se relaciona con la elevada reabsorción del sodio en los túbulos renales. Si el flujo renal y la FG se reducen y se filtra menos sodio, se reabsorbe menos sodio y se consume menos oxígeno
Determinantes del flujo sanguíneo renal El flujo sanguíneo renal está determinado por el gradiente de presión a través de los vasos renales (la diferencia entre las presiones hidrostáticas en la arteria renal y en la vena renal), dividido entre la resistencia vascular total renal
El flujo sanguíneo en los vasos rectos de la médula renal es muy bajo comparado con el flujo en la corteza renal La parte externa del riñón, la corteza renal, recibe la mayor parte del flujo sanguíneo renal. El flujo sanguíneo en la médula renal supone solo el 1-2% del flujo sanguíneo renal total. El flujo en la médula renal procede de una porción especializada del sistema capilar peritubular llamada vasos rectos. Estos vasos descienden hasta la médula paralelos a las asas de Henle y después vuelven de nuevo junto a las asas de Henle hasta la corteza antes de vaciarse en el sistema venoso
Control fisiológico de la filtración glomerular y del flujo sanguíneo renal
La intensa activación del sistema nervioso simpático reduce la FG Casi todos los vasos sanguíneos de los riñones, incluidas las arteriolas aferentes y eferentes, están muy inervados por fibras nerviosas simpáticas. La fuerte activación de los nervios simpáticos renales puede contraer las arteriolas renales y reducir el flujo sanguíneo renal y la FG. La estimulación moderada o leve ejerce poca influencia sobre el flujo sanguíneo renal y la FG
La noradrenalina, la adrenalina y la endotelina contraen los vasos sanguíneos renales y reducen la FG Las hormonas que constriñen las arteriolas aferentes y eferentes, lo que reduce la FG y el flujo sanguíneo renal, son la noradrenalina y la adrenalina liberadas por la médula suprarrenal. Las concentraciones sanguíneas de estas hormonas van generalmente paralelas a la actividad del sistema nervioso simpático; luego la noradrenalina y la adrenalina ejercen escasa influencia sobre la hemodinámica renal excepto en condiciones extremas, como una hemorragia grave. Otro vasoconstrictor, la endotelina, es un péptido que pueden liberar las células endoteliales vasculares lesionadas de los riñones, así como de otros tejidos.
La angiotensina II contrae preferentemente las arteriolas eferentes en la mayoría de los estados fisiológicos las arteriolas eferentes son altamente sensibles a la angiotensina II. Debido a que la angiotensina II contrae sobre todo las arteriolas eferentes en la mayoría de los estados fisiológicos, las concentraciones de angiotensina II aumentadas elevan la presión hidrostática glomerular mientras reducen el flujo sanguíneo renal. Debe tenerse en cuenta que la mayor formación de angiotensina II suele tener lugar en situaciones que se acompañan de una reducción de la presión arterial o una pérdida de volumen, que tienden a reducir la FG
Autorregulación de la FG y del flujo sanguíneo renal
Los mecanismos de retroalimentación intrínsecos de los riñones mantienen normalmente el flujo sanguíneo renal y la FG relativamente constantes, a pesar de cambios acentuados en la presión arterial sistémica.
Retroalimentación tubuloglomerular y autorregulación de la FG Los riñones tienen un mecanismo especial de retroalimentación que acopla los cambios en la concentración de cloruro de sodio en la mácula densa al control de la resistencia arteriolar renal y la autorregulación de la FG. Esta retroalimentación ayuda a asegurar una llegada relativamente constante de cloruro de sodio al túbulo distal y ayuda a evitar las fluctuaciones falsas en la excreción renal que de otro modo tendrían lugar. En muchas circunstancias, esta retroalimentación autorregula el flujo sanguíneo renal y la FG en paralelo
El mecanismo de retroalimentación tubuloglomerular tiene dos componentes que actúan juntos en el control de la FG: 1) un mecanismo de retroalimentación arteriolar aferente, y 2) un mecanismo de retroalimentación arteriolar eferente. Estos mecanismos de retroalimentación dependen de disposiciones anatómicas especiales del complejo yuxtaglomerular
Los estudios experimentales hacen pensar que la reducción de la FG disminuye la velocidad del flujo que llega al asa de Henle, lo que aumenta la reabsorción del porcentaje de iones sodio y cloro suministrados a la rama ascendente del asa de Henle, hecho que disminuye la concentración de cloruro de sodio en las células de la mácula densa. Esta reducción de la concentración de cloruro de sodio inicia una señal que parte de la mácula densa y tiene dos efectos: 1) reduce la resistencia al flujo sanguíneo en las arteriolas aferentes, lo que eleva la presión hidrostática glomerular y ayuda a normalizar la FG, y 2) aumenta la liberación de renina en las células yuxtaglomerulares de las arteriolas aferente y eferente, que son los principales reservorios de renina
Autorregulación miógena del flujo sanguíneo renal y de la FG Otro mecanismo que contribuye al mantenimiento del flujo sanguíneo renal y de la FG relativamente constantes es la capacidad de cada vaso sanguíneo de resistirse al estiramiento durante el aumento de la presión arterial, un fenómeno denominado mecanismo miógeno. Los estudios realizados en vasos individuales (sobre todo en arteriolas pequeñas) de todo el cuerpo han demostrado que responden a un aumento de la tensión o un estiramiento de la pared con una contracción del músculo liso vascular.
La mayoría de las personas saben que los riñones tienen una función importante: eliminar del cuerpo los materiales de desecho que se han ingerido o que ha producido el metabolismo. Una segunda función que es especialmente crítica es controlar el volumen y la composición de los electrólitos de los líquidos corporales. En lo que respecta al agua y casi todos los electrólitos del cuerpo, el equilibrio entre los ingresos (debidos a la ingestión y a la producción metabólica) y las salidas (debidas a la excreción o al consumo metabólico) lo mantienen en gran medida los riñones. Esta función reguladora de los riñones mantiene el ambiente interno estable necesario para que las células desempeñen sus diversas actividades
los riñones ejercen numerosas funciones homeostáticas, entre ellas las siguientes: • Excreción de productos metabólicos de desecho y sustancias químicas extrañas. • Regulación de los equilibrios hídrico y electrolítico. • Regulación de la osmolalidad del líquido corporal y de las concentraciones de electrólitos. • Regulación de la presión arterial. • Regulación del equilibrio acidobásico. • Regulación de la producción de eritrocitos. • Secreción, metabolismo y excreción de hormonas. • Gluconeogenia
Anatomía fisiológica de los riñones Organización general de los riñones y de la vía urinaria
Los riñones se disponen en la pared posterior del abdomen, fuera de la cavidad peritoneal. Cada riñón de un ser humano adulto normal pesa unos 150 g y tiene el tamaño aproximado de un puño cerrado. La cara medial de cada riñón contiene una región con una muesca, llamada hilio, por la que pasan la arteria y vena renales, los linfáticos, la inervación y el uréter, que transporta la orina final desde el riñón hasta la vejiga, donde se almacena hasta que esta se vacía. El riñón está rodeado de una cápsula fibrosa y tensa que protege sus delicadas estructuras internas.
Si se cortan los riñones de arriba abajo, las dos regiones principales que pueden verse son la corteza externa y las regiones internas de la médula. La médula se divide en 8-10 masas de tejido en forma de cono llamadas pirámides renales. La base de cada pirámide se origina en el borde entre la corteza y la médula y termina en la papila, que se proyecta en el espacio de la pelvis renal, una continuación en forma de abanico de la porción superior del uréter. El borde externo de la pelvis se divide en bolsas abiertas, llamadas cálices mayores, que se extienden hacia abajo y se dividen en los cálices menores, que recogen la orina de los túbulos de cada papila.
Irrigación renal El riego sanguíneo de los dos riñones es normalmente de alrededor del 22% del gasto cardíaco, o 1.100 ml/min. La arteria renal entra en el riñón a través del hilio y después se ramifica progresivamente hasta formar las arterias interlobulares, las arterias arciformes, las arterias interlobulillares (también denominadas arterias radiales) y las arteriolas aferentes, que acaban en los capilares glomerulares, donde se filtran grandes cantidades de líquido y solutos (excepto las proteínas plasmáticas) para comenzar la formación de orina (fig. 26-3). Los extremos distales de los capilares de cada glomérulo coalescen hasta formar la arteriola eferente, que llega a la segunda red capilar, los capilares peritubulares, que rodean a los túbulos renales
La nefrona es la unidad funcional del riñón
Cada nefrona contiene: 1) un penacho de capilares glomerulares llamado glomérulo, por el que se filtran grandes cantidades de líquido desde la sangre, y 2) un túbulo largo en el que el líquido filtrado se convierte en orina en su camino a la pelvis del riñón. El glomérulo contiene una red de capilares glomerulares que se ramifican y anastomosan y que, comparados con otros capilares, tienen una presión hidrostática alta (de unos 60 mmHg). Los capilares glomerulares están revestidos de células epiteliales y todo el glomérulo está cubierto por la cápsula de Bowman. El líquido filtrado desde los capilares glomerulares circula hacia la cápsula de Bowman y después al túbulo proximal, que se encuentra en la corteza del riñón. Desde el túbulo proximal, el líquido fluye hacia el asa de Henle, que desciende hasta la médula renal. Cada asa consta de una rama descendente y otra ascendente. Las paredes de la rama descendente y el segmento inferior de la rama ascendente son muy finas y de ahí que se denominen segmento fino del asa de Henle. Después de que la rama ascendente del asa ha vuelto a la corteza, su pared se engruesa mucho y se denomina segmento grueso del asa ascendente.
Diferencias regionales en la estructura de la nefrona: nefronas corticales y yuxtamedulares Aunque cada nefrona tiene todos los componentes descritos, hay algunas diferencias dependiendo de la profundidad a la que se encuentre la nefrona dentro de la masa renal. Las nefronas que tienen glomérulos localizados en la corteza externa se denominan nefronas corticales; tienen asas de Henle cortas que penetran solo una distancia corta en la médula
Alrededor del 20-30% de las nefronas tienen glomérulos que se disponen en la profundidad de la corteza renal cerca de la médula y se denominan nefronas yuxtamedulares. Estas nefronas tienen asas de Henle grandes que discurren hasta la médula, en algunos casos con un recorrido completamente intramedular hasta desembocar en las papilas renales. Las estructuras vasculares que irrigan las nefronas yuxtamedulares también difieren de las que irrigan las nefronas corticales. En las nefronas corticales todo el sistema tubular está rodeado de una red extensa de capilares peritubulares. En las nefronas yuxtamedulares, las arteriolas eferentes largas se extienden desde los glomérulos hasta la médula externa y después se dividen en capilares peritubulares especializados, llamados vasos rectos, que se extienden hasta la médula al lado de las asas de Henle
Micción
La micción es el proceso mediante el cual la vejiga urinaria se vacía cuando está llena. Este proceso se realiza en dos pasos. Primero, la vejiga se llena progresivamente hasta que la tensión en sus paredes aumenta por encima de un umbral. Esta tensión desencadena el segundo paso, que es un reflejo nervioso, llamado reflejo miccional, que vacía la vejiga o, si esto falla, provoca al menos un deseo de orinar. Aunque el reflejo miccional es un reflejo medular autónomo, centros presentes en la corteza cerebral o en el tronco del encéfalo pueden inhibirlo o facilitarlo.
Anatomía fisiológica de la vejiga La vejiga urinaria, que se muestra en la, es una cámara de músculo liso compuesta de dos partes principales: 1) el cuerpo, que es la principal parte de la vejiga en la que se acumula la orina, y 2) el cuello, que es una extensión en forma de abanico del cuerpo, que pasa en sentido inferior y anterior hasta el triángulo urogenital y se conecta con la uretra. La parte inferior del cuello de la vejiga también se llama uretra posterior por su relación con la uretra
El músculo liso de la vejiga se llama músculo detrusor. Sus fibras musculares se extienden en todas las direcciones y, cuando se contraen, pueden aumentar la presión en la vejiga hasta 40-60 mmHg. Luego la contracción del músculo detrusor es un paso importante en el vaciamiento de la vejiga. Las célula musculares lisas del músculo detrusor se fusionan entre sí de manera que existen vías eléctricas de baja resistencia de una célula muscular a otra. De este modo un potencial de acción puede propagarse a través del músculo detrusor, desde una célula muscular a la siguiente, para provocar la contracción de toda la vejiga a la vez.
En la pared posterior de la vejiga, inmediatamente por encima del cuello de la vejiga, hay una pequeña zona triangular llamada trígono. En la parte más inferior del vértice del trígono, el cuello de la vejiga se abre en la uretra posterior, y los dos uréteres entran en la vejiga en los ángulos más superiores del trígono. El trígono puede identificarse por el hecho de que su mucosa, el recubrimiento interno de la vejiga, es lisa, al contrario que el resto de la mucosa vesical, que está plegada y forma arrugas. Cada uréter, en su entrada en la vejiga, discurre en sentido oblicuo a través del músculo detrusor y después pasa otros 1-2 cm por debajo de la mucosa vesical antes de vaciarse en la vejiga
Inervación de la vejiga La principal inervación nerviosa de la vejiga es a través de los nervios pélvicos, que conectan con la médula espinal a través del plexo sacro, sobre todo los segmentos S2 y S3. En los nervios pélvicos discurren fibras nerviosas sensitivas y motoras. Las fibras sensitivas detectan el grado de distensión de la pared de la vejiga. Las señales de distensión de la uretra posterior son especialmente fuertes y son responsables sobre todo de iniciar los reflejos que provocan el vaciado de la vejiga.
Filtración, reabsorción y secreción de diferentes sustancias En general, la reabsorción tubular es cuantitativamente más importante que la secreción tubular en la formación de la orina, pero la secreción es importante para determinar las cantidades de iones potasio e hidrógeno y algunas otras sustancias que se excretan por la orina. La mayoría de las sustancias que deben eliminarse de la sangre, en especial los productos finales del metabolismo, como la urea, la creatinina, el ácido úrico y los uratos, se reabsorben mal y por ello se excretan en grandes cantidades en la orina. Ciertas sustancias extrañas y fármacos se reabsorben mal pero, además, se secretan desde la sangre a los túbulos, de manera que su excreción es alta. Por el contrario, los electrólitos, como los iones cloro, sodio y bicarbonato, se reabsorben mucho, de manera que solo se detectan en la orina pequeñas cantidades. Ciertas sustancias nutritivas, como los aminoácidos y la glucosa, se reabsorben completamente de los túbulos y no aparecen en la orina, aunque los capilares glomerulares filtren grandes cantidades. Cada uno de los procesos (filtración glomerular, reabsorción y secreción tubular) está regulado de acuerdo con las necesidades del cuerpo
La ingestión y la pérdida de líquido están equilibradas durante las situaciones estables
Resulta llamativa la relativa constancia de los líquidos corporales ya que hay un intercambio continuo de líquido y solutos con el ambiente externo, así como dentro de los diferentes compartimientos del cuerpo.
Ingestión diaria de agua El agua ingresa en el cuerpo a través de dos fuentes principales: 1) se ingiere en forma de líquidos o de agua de los alimentos, que juntos suponen alrededor de 2.100 ml/día de líquidos corporales, y 2) se sintetiza en el cuerpo por la oxidación de los hidratos de carbono, en una cantidad de unos 200 ml/día. Estos mecanismos proporcionan un ingreso total de agua de unos 2.300 ml/dia. Sin embargo, la ingestión de agua es muy variable entre las diferentes personas e incluso en la misma persona en diferentes días en función del clima, los hábitos e incluso el grado de actividad física
Pérdida diaria de agua corporal
Pérdida insensible de agua Parte de las pérdidas de agua no puede regularse de manera precisa. Por ejemplo, los seres humanos experimentan una pérdida continua de agua por evaporación de las vías aéreas y por difusión a través de la piel, y en conjunto son responsables de alrededor de 700 ml/día de pérdida de agua en condiciones normales. A esto se le denomina pérdida insensible de agua porque no somos conscientes de ella, aunque se produzca continuamente en todos los seres humanos vivos.
Pérdida de líquido en el sudor La cantidad de agua perdida por el sudor es muy variable dependiendo de la actividad física y de la temperatura ambiental. El volumen de sudor es normalmente de unos 100 ml/día, pero en un clima muy cálido o durante el ejercicio intenso, la pérdida de líquidos en el sudor aumenta en ocasiones a 1-2 l/h. Esta pérdida vaciaría rápidamente los líquidos corporales si la ingestión no aumentara mediante la activación del mecanismo de la sed
Pérdida de agua en las heces Solo se pierde normalmente una pequeña cantidad de agua (100 ml/día) en las heces. Esta pérdida puede aumentar a varios litros al día en personas con diarrea intensa.
Pérdida de agua por los riñones El resto del agua perdida se excreta en la orina por los riñones. Múltiples mecanismos controlan la intensidad de la producción de orina. De hecho, el medio más importante por el que el cuerpo mantiene un equilibrio entre los ingresos y las pérdidas, así como el equilibrio entre el ingreso y la salida de la mayoría de los electrólitos en el cuerpo, es controlando la intensidad con la que los riñones excretan estas sustancias.
Compartimientos del líquido corporal
El líquido corporal total se distribuye sobre todo entre dos compartimientos: el líquido extracelular y el líquido intracelular. El líquido extracelular se divide en el líquido intersticial y el plasma sanguíneo. Existe otro pequeño compartimiento de líquido que se denomina líquido transcelular. Este compartimiento comprende el líquido de los espacios sinovial, peritoneal, pericárdico e intracelular, así como el líquido cefalorraquídeo; suele considerarse un tipo especializado de líquido extracelular, aunque en algunos casos su composición puede diferir de forma acentuada de la del plasma o de la del líquido intersticial. Todos los líquidos transcelulares constituyen alrededor de 1 a 2 l. En un hombre adulto de 70 kg, el agua corporal total representa alrededor del 60% del peso corporal (unos 42 l). Este porcentaje depende de la edad, el sexo y el grado de obesidad.
Compartimiento del líquido intracelular Unos 28 de los 42 l de líquido corporal están contenidos de los 100 billones de células y se les denomina en conjunto líquido intracelular. Por tanto, el líquido intracelular constituye alrededor del 40% del peso corporal total en una persona «media». El líquido de cada célula contiene una mezcla individual de diferentes constituyentes, pero las concentraciones de estas sustancias son similares de una célula a otra
Compartimiento del líquido extracelular Todos los líquidos del exterior de las células se denominan en conjunto líquido extracelular y constituyen alrededor del 20% del peso corporal, o unos 14 l en un hombre adulto de 70 kg. Los dos compartimientos más grandes del líquido extracelular son el líquido intersticial, que supone hasta más de tres cuartas partes (11 l) del líquido extracelular, y el plasma, que supone casi una cuarta parte del líquido extracelular o unos 3L.
Volumen sanguíneo La sangre contiene líquido extracelular (el líquido del plasma) y líquido intracelular (el líquido de los eritrocitos). Sin embargo, la sangre se considera un compartimiento líquido separado porque está contenida en su propia cámara, el aparato circulatorio. El volumen sanguíneo es especialmente importante en el control de la dinámica cardiovascular. El volumen sanguíneo medio de los adultos es de alrededor del 7% del peso corporal (unos 5 l). Alrededor del 60% de la sangre es plasma y el 40% son eritrocitos, pero estos porcentajes pueden variar considerablemente en diferentes personas dependiendo del sexo, el peso y otros factores.
Constituyentes de los líquidos extracelular e intracelular
La composición iónica del plasma y del líquido intersticial es similar Debido a que el plasma y el líquido intersticial están separados solo por membranas capilares muy permeables, su composición iónica es similar. La diferencia más importante entre estos dos compartimientos es la mayor concentración de proteínas en el plasma; debido a que los capilares tienen una permeabilidad baja a las proteínas plasmáticas, solo pequeñas cantidades de proteínas pasan a los espacios intersticiales en la mayoría de los tejidos
Constituyentes del líquido intracelular El líquido intracelular está separado del líquido extracelular por una membrana celular que es muy permeable al agua, pero no a la mayoría de los electrólitos del cuerpo. Al contrario que el líquido extracelular, el líquido intracelular contiene solo mínimas cantidades de iones sodio y cloro y casi ningún ion calcio. En cambio, contiene grandes cantidades de iones potasio y fosfato más cantidades moderadas de iones magnesio y sulfato, todos los cuales están en concentraciones bajas en el líquido extracelular. Además, las células contienen grandes cantidades de proteínas, casi cuatro veces más que en el plasma.
Regulación del intercambio de líquido y del equilibrio osmótico entre los líquidos intracelular y extracelular
las cantidades relativas de líquido extracelular distribuidas entre los espacios plasmático e intersticial están determinadas sobre todo por el equilibrio entre las fuerzas hidrostática y coloidosmótica a través de las membranas capilares. La distribución del líquido entre los compartimientos intracelular y extracelular, en cambio, está determinada sobre todo por el efecto osmótico de los solutos más pequeños (en especial el sodio, el cloro y otros electrólitos) que actúan a través de la membrana celular
Principios básicos de la ósmosis y la presión osmótica Debido a que las membranas celulares son relativamente impermeables a la mayoría de los solutos pero muy permeables al agua (es decir, son permeables selectivamente), donde quiera que haya una mayor concentración de soluto a un lado de la membrana celular, el agua se difundirá a través de la membrana hacia la región de mayor concentración de soluto
Osmolalidad y osmolaridad La concentración osmolal de una solución se denomina osmolalidad cuando la concentración se expresa en osmoles por kilogramo de agua; se llama osmolaridad cuando se expresa en osmoles por litro de solución.
Osmolaridad de los líquidos corporales alrededor del 80% de la osmolaridad total del líquido intersticial y del plasma se debe a los iones sodio y cloro, mientras que en el líquido intracelular, casi la mitad de la osmolaridad se debe a los iones potasio, y el resto se divide entre muchas otras sustancias intracelulares.
Si una célula se coloca en una solución de solutos no difusibles con una osmolaridad de 282 mOsm/l, las células no se encogerán ni hincharán porque la concentración de agua en los líquidos extracelular e intracelular es igual y los solutos no pueden entrar ni salir de la célula. Se dice que este tipo de solución es isotónica porque ni encoge ni hincha las células.
Si se coloca una célula en una solución hipotónica que tiene una menor concentración de solutos no difusibles (menos de 282 mOsm/l), el agua se difundirá al interior de la célula y la hinchará; el agua continuará difundiendo al interior de la célula diluyendo el líquido intracelular mientras concentra el líquido extracelular hasta que ambas soluciones tengan la misma osmolaridad. Las soluciones de cloruro de sodio con una concentración menor de un 0,9% son hipotónicas e hincharán a la célula. Si se coloca una célula en una solución hipertónica con una solución mayor de solutos no difusibles, el agua saldrá de la célula hacia el líquido extracelular concentrando el líquido intracelular y diluyendo el líquido extracelular. En este caso la célula se contraerá hasta que las dos concentraciones se igualen. Las soluciones de cloruro de sodio mayores del 0,9% son hipertónicas
Efecto de la adición de una solución salina al líquido extracelular
Si se añade una solución salina isotónica al compartimiento líquido extracelular, la osmolaridad del líquido extracelular no cambia y no se produce ninguna ósmosis a través de las membranas celulares. El único efecto es un aumento del volumen de líquido extracelular
Si se añade una solución hipertónica al líquido extracelular, la osmolaridad extracelular aumenta y provoca la ósmosis del agua fuera de las células hacia el compartimiento extracelular. De nuevo, casi todo el cloruro de sodio añadido permanece en el compartimiento extracelular y el líquido difunde desde las células hacia el espacio extracelular hasta conseguir el equilibrio osmótico. El efecto neto es un aumento del volumen extracelular (mayor del volumen de líquido añadido), una reducción del volumen intracelular y un aumento de la osmolaridad en los dos compartimientos. Si se añade una solución hipotónica al líquido extracelular, la osmolaridad disminuye y parte del agua extracelular difunde al interior de las células hasta que los compartimientos extracelular e intracelular tienen la misma osmolaridad. Los volúmenes extracelular e intracelular aumentan al añadir líquido hipotónico, aunque el volumen intracelular lo hace en mayor grado
Edema: exceso de líquido en los tejidos
El edema se refiere a la presencia de un exceso de líquido en los tejidos corporales. En la mayoría de los casos el edema aparece sobre todo en el compartimiento de líquido extracelular, pero puede afectar también al líquido intracelular.
Edema intracelular Tres procesos causan especialmente tumefacción o edema intracelular: 1) la hiponatremia, según se ha comentado anteriormente; 2) la depresión de los sistemas metabólicos de los tejidos, y 3) la falta de una nutrición celular adecuada
Edema extracelular El edema extracelular se produce cuando se acumula un exceso de líquido en los espacios extracelulares. Hay dos causas generales de edema extracelular: 1) la fuga anormal de líquido del plasma hacia los espacios intersticiales a través de los capilares, y 2) la imposibilidad de los linfáticos de devolver el líquido a la sangre desde el intersticio, lo que a menudo se conoce por linfedema. La causa clínica más común de la acumulación intersticial de líquido es la filtración capilar excesiva de líquido.
Cuando el impulso cardíaco atraviesa el corazón, la corriente eléctrica también se propaga desde el corazón hacia los tejidos adyacentes que lo rodean. Una pequeña parte de la corriente se propaga hacia la superficie corporal. Si se colocan electrodos en la piel en lados opuestos del corazón se pueden registrar los potenciales eléctricos que se generan por la corriente; el registro se conoce como electrocardiograma (ECG).
CARACTERÍSTICAS DEL ELECTROCARDIOGRAMA NORMAL
El ECG normal está formado por una onda P, un complejo QRS y una onda T. Con frecuencia, aunque no siempre, el complejo QRS está formado por tres ondas separadas: la onda Q, la onda R y la onda S.
La onda P está producida por los potenciales eléctricos que se generan cuando se despolarizan las aurículas antes del comienzo de la contracción auricular. El complejo QRS está formado por los potenciales que se generan cuando se despolarizan los ventrículos antes de su contracción, es decir, a medida que la onda de despolarización se propaga por los ventrículos. Por tanto, tanto la onda P como los componentes del complejo QRS son las ondas de despolarización.
La onda T está producida por los potenciales que se generan cuando los ventrículos se recuperan del estado de despolarización. Este proceso normalmente aparece en el músculo ventricular entre 0,25 y 0,35 s después de la despolarización. La onda T se conoce como onda de repolarización.
Así, el ECG está formado por ondas tanto de despolarización como de repolarización. Los principios de la despolarización y de la repolarización se analizan en el capítulo 5. La distinción entre ondas de despolarización y ondas de repolarización es tan importante en electrocardiografía que requiere una aclaración adicional.
ONDAS DE DESPOLARIZACIÓN FRENTE A ONDAS DE REPOLARIZACIÓN
Durante la despolarización el potencial negativo normal del interior de la fibra se invierte y se hace ligeramente positivo en el interior y negativo en el exterior.
Relación del potencial de acción monofásico del músculo ventricular con las ondas QRS y T del electrocardiograma estándar
El potencial de acción monofásico del músculo ventricular, que se ha analizado en el capítulo 10, normalmente dura entre 0,25 y 0,35 s. El potencial de acción monofásico registrado con un microelectrodo insertado en el interior de una fibra muscular ventricular única. El ascenso de este potencial de acción está producido por la despolarización, y la vuelta del potencial al nivel basal está producida por la repolarización.
En un registro simultáneo del ECG de un ventrículo. Las ondas QRS aparecen al principio del potencial de acción monofásico y la onda T aparece al final. Obsérvese específicamente que no se registra ningún potencial en el ECG cuando el músculo ventricular está completamente polarizado o completamente despolarizado. Solo cuando el músculo está parcialmente polarizado o parcialmente despolarizado hay flujo de corriente desde una parte de los ventrículos hacia la otra, y por tanto la corriente también fluye hacia la superficie del cuerpo para generar el ECG.
RELACIÓN DE LA CONTRACCIÓN AURICULAR Y VENTRICULAR CON LAS ONDAS DEL ELECTROCARDIOGRAMA
Antes de que se pueda producir la contracción del músculo, la despolarización se debe propagar por todo el músculo para iniciar los procesos químicos de la contracción. La onda P se produce al comienzo de la contracción de las aurículas y el complejo QRS de ondas se produce al comienzo de la contracción de los ventrículos. Los ventrículos siguen contraídos hasta después de que se haya producido la repolarización, es decir, hasta después del final de la onda T.
Las aurículas se repolarizan aproximadamente 0,15 a 0,2 s después de la finalización de la onda P, lo que coincide aproximadamente con el momento en el que se registra el complejo QRS en el ECG. Por tanto, la onda de repolarización auricular, conocida como onda T auricular, habitualmente está oscurecida por el complejo QRS, que es mucho mayor. Por este motivo raras veces se observa la onda T auricular en el ECG.
La onda de repolarización ventricular es la onda T del ECG normal. Habitualmente el músculo ventricular comienza a repolarizarse en algunas fibras aproximadamente 0,2 s después del comienzo de la onda de despolarización (el complejo QRS), pero en muchas otras fibras tarda hasta 0,35 s. Así, el proceso de repolarización ventricular se extiende a lo largo de un período prolongado, de aproximadamente 0,15 s. Por este motivo la onda T del ECG normal es una onda prolongada, aunque el voltaje de la onda T es mucho menor que el voltaje del complejo QRS, en parte debido a esta duración prolongada.
Calibración del voltaje y el tiempo del electrocardiograma
Todos los registros de los ECG se hacen con líneas de calibración adecuadas sobre el papel de registro. Estas líneas de calibración pueden estar ya señaladas en el papel, como ocurre cuando se utiliza un registrador de pluma, o se registran en el papel al mismo tiempo que se registra el ECG, como en los tipos fotográficos de electrocardiógrafos.
Las líneas de calibración horizontal están dispuestas de modo que 10 de las divisiones de las líneas pequeñas hacia arriba o hacia abajo en el ECG estándar representan 1 mV, con la positividad hacia arriba y la negatividad hacia abajo.
Las líneas verticales del ECG son las líneas de calibración del tiempo. Un ECG típico se realiza a una velocidad de papel de 25 mm/s, aunque en ocasiones se emplean velocidades más rápidas. Por tanto, cada 25 mm en dirección horizontal corresponden a 1 s y cada segmento de 5 mm, indicado por las líneas verticales oscuras, representa 0,2 s. Después los intervalos de 0,2 s están divididos en cinco intervalos más pequeños por líneas finas, cada una de las cuales representa 0,04 s.
Voltajes normales en el electrocardiograma
Los voltajes de las ondas que se registran en el ECG normal dependen de la manera en la que se aplican los electrodos a la superficie del cuerpo y de la proximidad de los electrodos al corazón. Cuando un electrodo está colocado directamente sobre los ventrículos y un segundo electrodo está localizado en otra localización del cuerpo alejada del corazón, el voltaje del complejo QRS puede ser de hasta 3 a 4 mV.
Incluso este voltaje es pequeño en comparación con el potencial de acción monofásico de 110 mV que se registra directamente en la membrana del músculo cardíaco. Cuando los ECG se registran con electrodos en los dos brazos o en un brazo y una pierna, el voltaje en el complejo QRS habitualmente es de 1 a 1,5 mV desde el punto más elevado de la onda R hasta el punto más profundo de la onda S; el voltaje de la onda P está entre 0,1 y 0,3 mV, y el de la onda T está entre 0,2 y 0,3 mV.
Intervalo P-Q o P-R
El tiempo que transcurre entre el comienzo de la onda P y el comienzo del complejo QRS es el intervalo que hay entre el inicio de la excitación eléctrica de las aurículas y el inicio de la excitación de los ventrículos. Este período se denomina intervalo P-Q. El intervalo P-Q normal es de aproximadamente 0,16 s. (Con frecuencia este intervalo se denomina intervalo P-R porque es probable que no haya onda Q.)
Intervalo Q-T
La contracción del ventrículo dura casi desde el comienzo de la onda Q (onda R si no hay onda Q) hasta el final de la onda T. Este intervalo se denomina intervalo Q-T y habitualmente es de aproximadamente 0,35 s.
Determinación de la frecuencia del latido cardíaco a partir del electrocardiograma
La frecuencia del latido cardíaco se puede determinar fácilmente a partir del ECG porque la frecuencia cardíaca es el recíproco del intervalo de tiempo entre dos latidos cardíacos sucesivos. Si el intervalo entre dos latidos, que se determina a partir de las líneas de calibración del tiempo, es de 1 s, la frecuencia cardíaca es de 60 latidos/min. El intervalo normal entre dos complejos QRS sucesivos en una persona adulta es de aproximadamente 0,83 s, lo que corresponde a una frecuencia cardíaca de 60/0,83 veces por minuto, o 72 latidos/min.
FLUJO DE CORRIENTE ALREDEDOR DEL CORAZÓN DURANTE EL CICLO CARDÍACO
REGISTRO DE POTENCIALES ELÉCTRICOS A PARTIR DE UNA MASA PARCIALMENTE DESPOLARIZADA DE MÚSCULO CARDÍACO SINCITIAL
Una masa sincitial de músculo cardíaco que ha sido estimulada en su punto más central. Antes de la estimulación, el exterior de todas las células musculares era positivo y el interior negativo. Por los motivos en el análisis de los potenciales de membrana, tan pronto como se despolariza una zona del sincitio cardíaco se produce la salida de cargas negativas hacia el exterior de las fibras musculares despolarizadas, haciendo que esta parte de la superficie sea electronegativa. El resto de la superficie del corazón, que sigue polarizada, está representado por los signos positivos. Por tanto, un medidor conectado con el terminal negativo en la zona de despolarización y el terminal positivo en una de las zonas que todavía están polarizadas, como se muestra a la derecha de la figura, registra un valor positivo.
También se presentan otras dos disposiciones de los electrodos y lecturas de los medidores. Estas disposiciones y lecturas se deben estudiar cuidadosamente, y el lector debe ser capaz de explicar las causas de las respectivas lecturas de los medidores. Como la despolarización se propaga por el corazón en todas las direcciones, las diferencias de potencial persisten solo durante algunas milésimas de segundo, y las mediciones del voltaje real solo se pueden realizar con un aparato de registro de alta velocidad.
FLUJO DE CORRIENTES ELÉCTRICAS EN EL TÓRAX ALREDEDOR DEL CORAZÓN
El músculo ventricular situado en el interior del tórax. Incluso los pulmones, aunque están llenos de aire en su mayor parte, conducen la electricidad en una magnitud sorprendente, y los líquidos de los demás tejidos que rodean el corazón conducen la electricidad incluso con más facilidad. Por tanto, el corazón realmente está suspendido en un medio conductor. Cuando una porción de los ventrículos se despolariza y, por tanto, se hace electronegativa en relación con el resto, la corriente eléctrica fluye desde la zona despolarizada hacia la zona polarizada en rutas sinuosas largas.
Del análisis del sistema de Purkinje se debe recordar que la primera zona de los ventrículos a la que llega el impulso cardíaco es el tabique, y poco después se propaga a la superficie interna del resto de la masa de los ventrículos, por las zonas rojas y los signos negativo. Este proceso hace que las zonas internas de los ventrículos sean electronegativas y que las paredes externas de los ventrículos sean electropositivas, de modo que la corriente eléctrica fluye a través de los líquidos que rodean los ventrículos en trayectos elípticos. Si se realiza el promedio algebraico de todas las líneas de flujo de corriente (las líneas elípticas) se encuentra que el flujo medio de corriente tiene negatividad hacia la base del corazón y positividad hacia la punta. Durante la mayor parte del resto del proceso de despolarización la corriente también sigue fluyendo en esta misma dirección, mientras que la despolarización se propaga desde la superficie endocárdica hacia el exterior a través de la masa del músculo ventricular. Después, inmediatamente antes de que la despolarización haya completado su trayecto a través de los ventrículos, la dirección media del flujo de corriente se invierte durante aproximadamente 0,01 s, fluyendo desde la punta ventricular hacia la base, porque la última parte del corazón que se despolariza son las paredes externas de los ventrículos cerca de la base del corazón.
Así, en los ventrículos del corazón normal la corriente fluye desde las zonas negativas a las positivas principalmente en una dirección que va desde la base del corazón hacia la punta durante casi todo el ciclo de despolarización, excepto al final. Si se conecta un medidor a los electrodos de la superficie del cuerpo, el electrodo más próximo a la base será negativo, mientras que el electrodo más próximo a la punta será positivo, y el medidor de registro mostrará un registro positivo en el ECG.
DERIVACIONES ELECTROCARDIOGRÁFICAS
TRES DERIVACIONES BIPOLARES DE LAS EXTREMIDADES
Las conexiones eléctricas entre las extremidades del paciente y el electrocardiógrafo para registrar ECG de las denominadas derivaciones bipolares estándar de las extremidades. El término «bipolar» significa que el electrocardiograma se registra a partir de dos electrodos que están localizados en lados diferentes del corazón, en este caso en las extremidades. Así, una «derivación» no es un único cable que procede del cuerpo, sino una combinación de dos cables y sus electrodos para formar un circuito completo entre el cuerpo y el electrocardiógrafo. En cada uno de los casos el electrocardiógrafo se representa en el diagrama mediante un medidor eléctrico, aunque el electrocardiógrafo real es un sistema informático de alta velocidad con una pantalla electrónica.
Derivación I
Cuando se registra la derivación I, el terminal negativo del electrocardiógrafo está conectado al brazo derecho y el terminal positivo al brazo izquierdo. Por tanto, cuando el punto en el que el brazo derecho se conecta con el tórax es electronegativo respecto al punto en el que se conecta el brazo izquierdo el electrocardiógrafo registra una señal positiva, es decir, por encima de la línea de voltaje cero del ECG. Cuando ocurre lo contrario el electrocardiógrafo registra una señal por debajo de la línea.
Derivación II
Para registrar la derivación II de las extremidades, el terminal negativo del electrocardiógrafo se conecta al brazo derecho y el terminal positivo a la pierna izquierda. Por tanto, cuando el brazo derecho es negativo respecto a la pierna izquierda, el
electrocardiógrafo registra una señal positiva.
Derivación III
Para registrar la derivación III de las extremidades, el terminal negativo del electrocardiógrafo se conecta al brazo izquierdo y el terminal positivo a la pierna izquierda. Esta configuración significa que el electrocardiógrafo registra una señal positiva cuando el brazo izquierdo es negativo respecto a la pierna izquierda.
Triángulo de Einthoven
En el cuerpo se dibuja un triángulo, denominado triángulo de Einthoven, alrededor de la zona del corazón. Este diagrama ilustra que los dos brazos y la pierna izquierda forman vértices de un triángulo que rodea el corazón. Los dos vértices de la parte superior del triángulo representan los puntos en los que los dos brazos se conectan eléctricamente a los líquidos que rodean el corazón y el vértice izquierdo es el punto en el que la pierna izquierda se conecta a los líquidos.
Ley de Einthoven
La ley de Einthoven afirma que, si los ECG se registran simultáneamente en las tres derivaciones de las extremidades, la suma de los potenciales registrados en las derivaciones I y III debe ser igual al potencial en la derivación II.
En otras palabras, si en cualquier momento dado se conocen los potenciales eléctricos de dos cualesquiera de las tres derivaciones electrocardiográficas bipolares de las extremidades, se puede determinar la tercera simplemente sumando las dos primeras. Ha de tenerse en cuenta, sin embargo, que se deben observar los signos positivos y negativos de las diferentes derivaciones cuando se haga esta suma.
Electrocardiogramas normales registrados en las tres derivaciones bipolares estándar de las extremidades
El registro de los ECG de las derivaciones I, II y III. Es evidente que los ECG de estas tres derivaciones son similares entre sí porque todos registran ondas P positivas y ondas T positivas, y la mayor parte del complejo QRS también es positiva en todos los ECG.
Cuando se analizan los tres ECG se puede demostrar, con mediciones cuidadosas y teniendo en cuenta las polaridades, que en cualquier momento dado la suma de los potenciales de las derivaciones I y III es igual al potencial de la derivación II, lo que ilustra la validez de la ley de Einthoven.
Como los registros de todas las derivaciones bipolares de las extremidades son similares entre sí, no importa mucho qué derivación se registra cuando se quieren diagnosticar diferentes arritmias cardíacas, porque el diagnóstico de las arritmias depende principalmente de las relaciones temporales entre las diferentes ondas del ciclo cardíaco. Sin embargo, cuando se desea diagnosticar la lesión del músculo ventricular o auricular o del sistema de conducción de Purkinje sí importa mucho qué derivaciones se registran, porque las alteraciones de la contracción del músculo cardíaco o de la conducción del impulso cardíaco modifican mucho los patrones de los ECG en algunas derivaciones, aunque pueden no afectar a otras. La interpretación electrocardiográfica de estos dos tipos de enfermedades (miopatías cardíacas y arritmias cardíacas)
DERIVACIONES DEL TÓRAX (DERIVACIONES PRECORDIALES)
Con frecuencia se registran ECG con un electrodo situado en la superficie anterior del tórax directamente sobre el corazón en uno de los puntos. Este electrodo se conecta al terminal positivo del electrocardiógrafo, y el electrodo negativo, denominado electrodo indiferente, se conecta a través de resistencias eléctricas iguales al brazo derecho, al brazo izquierdo y a la pierna izquierda al mismo tiempo. Habitualmente se registran seis derivaciones estándar del tórax, una cada vez, desde la pared torácica anterior, de modo que el electrodo del tórax se coloca secuencialmente en los seis puntos que se muestran en el diagrama. Los diferentes registros se conocen como derivaciones V1, V2, V3, V4, V5 y V6.
En las derivaciones V1 y V2 los registros QRS del corazón normal son principalmente negativos porque, como se muestra en la figura 11-8, el electrodo del tórax de estas derivaciones está más cerca de la base del corazón que de la punta, y la base del corazón está en la dirección de la electronegatividad durante la mayor parte del proceso de despolarización ventricular. Por el contrario, los complejos QRS de las derivaciones V4, V5 y V6 son principalmente positivos porque el electrodo del tórax de estas derivaciones está más cerca de la punta cardíaca, que está en la dirección de la electropositividad durante la mayor parte de la despolarización.
DERIVACIONES UNIPOLARES AMPLIADAS DE LAS EXTREMIDADES
Otro sistema de derivaciones que se utiliza mucho es la derivación unipolar ampliada de las extremidades. En este tipo de registro, dos de las extremidades se conectan mediante resistencias eléctricas al terminal negativo del electrocardiógrafo, y la tercera extremidad se conecta al terminal positivo. Cuando el terminal positivo está en el brazo derecho la derivación se conoce como derivación aVR, cuando está en el brazo izquierdo es la derivación aVL y cuando está en la pierna izquierda es la derivación aVF.
El corazón humano tiene un sistema especial para la autoexcitación rítmica y la contracción repetitiva aproximadamente 100.000 veces al día, o 3.000 millones de veces en una vida humana de duración media. Este impresionante logro es realizado por un sistema que:
Genera impulsos eléctricos rítmicos para iniciar la contracción rítmica del músculo cardíaco.
Conduce estos estímulos rápidamente por todo el corazón.
Cuando este sistema funciona normalmente, las aurículas se contraen aproximadamente 1/6 de segundo antes de la contracción ventricular, lo que permite el llenado de los ventrículos antes de que bombeen la sangre a través de los pulmones y de la circulación periférica. Este sistema también es importante porque permite que todas las porciones de los ventrículos se contraigan casi simultáneamente, lo que es esencial para una generación de presión más eficaz en las cavidades ventriculares.
Este sistema rítmico y de conducción del corazón se puede lesionar en las cardiopatías, especialmente en la isquemia de los tejidos cardíacos que se debe a una disminución del flujo sanguíneo coronario. La consecuencia es con frecuencia una alteración del ritmo cardíaco o una secuencia anormal de contracción de las cavidades cardíacas, con una posible alteración grave de la eficacia de la función de bomba del corazón, incluso hasta el grado de producir la muerte.
SISTEMA DE EXCITACIÓN ESPECIALIZADO Y DE CONDUCCIÓN DEL CORAZÓN
El sistema especializado de excitación y conducción del corazón que controla las contracciones cardíacas. El nódulo sinusal (también denominado nódulo sinoauricular o SA), en el que se genera el impulso rítmico normal; las vías internodulares que conducen impulsos desde el nódulo sinusal hasta el nódulo auriculoventricular (AV); el nódulo AV, en el cual los impulsos originados en las aurículas se retrasan antes de penetrar en los ventrículos; el haz AV, que conduce impulsos desde las aurículas hacia los ventrículos, y las ramas izquierda y derecha del haz de fibras de Purkinje, que conducen los impulsos cardíacos por todo el tejido de los ventrículos.
NÓDULO SINUSAL (SINOAURICULAR)
El nódulo sinusal (también denominado nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Está localizado en la pared posterolateral superior de la aurícula derecha, inmediatamente inferior y ligeramente lateral a la desembocadura de la vena cava superior. Las fibras de este nódulo casi no tienen filamentos musculares contráctiles y cada una de ellas tiene solo de 3 a 5 μm de diámetro, en contraste con un diámetro de 10 a 15 μm para las fibras musculares auriculares circundantes. Sin embargo, las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
RITMICIDAD ELÉCTRICA AUTOMÁTICA DE LAS FIBRAS SINUSALES
Algunas fibras cardíacas tienen la capacidad de autoexcitación, que es un proceso que puede producir descargas y contracciones rítmicas automáticas. Esta capacidad es especialmente cierta en el caso de las fibras del sistema especializado de conducción del corazón, entre ellas las fibras del nódulo sinusal. Por este motivo el nódulo sinusal habitualmente controla la frecuencia del latido de todo el corazón, como se analiza en detalle más adelante en este mismo capítulo. En primer lugar, se va a describir esta ritmicidad automática.
Mecanismo de la ritmicidad del nódulo sinusal. Los potenciales de acción registrados desde el interior de una fibra del nódulo sinusal durante tres latidos cardíacos y, a modo de comparación, un único potencial de acción de una fibra muscular ventricular. Entre descargas el «potencial de membrana en reposo» de la fibra del nódulo sinusal tiene una negatividad de aproximadamente –55 a –60 mV, en comparación con –85 a – 90 mV para la fibra muscular ventricular. La causa de esta menor negatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio, y las cargas positivas de los iones sodio y calcio que entran neutralizan parte de la negatividad intracelular.
La ritmicidad de las fibras del nódulo sinusal muestran de que el músculo cardíaco tiene tres tipos de canales iónicos de membrana que tienen funciones importantes en la generación de los cambios de voltaje en el potencial de acción. Los tipos son:
Los canales rápidos de sodio.
Los canales de calcio de tipo L (canales lentos de sodio-calcio)
Los canales de potasio.
La apertura de los canales rápidos de sodio durante algunas diezmilésimas de segundo es responsable de la rápida espiga ascendente del potencial de acción que se observa en el músculo ventricular, debido a la entrada rápida de iones sodio positivos hacia el interior de la fibra. Después, la «meseta» del potencial de acción ventricular está producida principalmente por la apertura más lenta de los canales lentos de sodio-calcio, que dura aproximadamente 0,3 s. Finalmente, la apertura de los canales de potasio permite la difusión de grandes cantidades de iones potasio positivos hacia el exterior a través de la membrana de la fibra y devuelve el potencial de membrana a su nivel de reposo.
Hay una diferencia en la función de estos canales en la fibra del nódulo sinusal porque el potencial «de reposo» es mucho menos negativo (de solo –55 mV en la fibra nodular, en lugar de los –90 mV de la fibra muscular ventricular). A este nivel de –55 mV, los canales rápidos de sodio principalmente ya se han «inactivado», lo que significa que han sido bloqueados. La causa de esto es que siempre que el potencial de membrana es menos negativo de aproximadamente –55 mV durante más de algunos milisegundos, las compuertas de inactivación del interior de la membrana celular que cierran los canales rápidos de sodio se cierran y permanecen de esta manera. Por tanto, solo se pueden abrir los canales lentos de sodio-calcio (es decir, se pueden «activar») y, por tanto, pueden producir el potencial de acción. En consecuencia, el potencial de acción del nódulo auricular se produce más lentamente que el potencial de acción del músculo ventricular. Además, después de la producción del potencial de acción, el regreso del potencial a su estado negativo también se produce lentamente, en lugar del regreso súbito que se produce en la fibra ventricular.
Autoexcitación de las fibras del nódulo sinusal
Debido a la elevada concentración de iones sodio en el líquido extracelular en el exterior de la fibra nodular, así como al número moderado de canales de sodio abiertos previamente, los iones sodio positivos del exterior de las fibras normalmente tienden a desplazarse hacia el interior. Por tanto, entre los latidos cardíacos, la entrada de iones sodio de carga positiva produce una elevación lenta del potencial de membrana en reposo en dirección positiva. Así, el potencial «en reposo» aumenta gradualmente y se hace menos negativo entre cada dos latidos sucesivos. Cuando el potencial alcanza un voltaje umbral de aproximadamente –40 mV, los canales de calcio de tipo L se «activan», produciendo de esta manera el potencial de acción. Por tanto, básicamente, la permeabilidad inherente de las fibras del nódulo sinusal a los iones sodio y calcio produce su autoexcitación.
Durante el transcurso del potencial de acción se producen dos fenómenos que impiden dicho estado de despolarización constante. Primero, los canales de calcio de tipo L se inactivan (es decir, se cierran) en un plazo de aproximadamente 100 a 150 ms después de su apertura, y segundo, aproximadamente al mismo tiempo se abren números muy elevados de canales de potasio. Por tanto, se interrumpe el flujo de entrada de iones positivos calcio y sodio a través de los canales de calcio de tipo L, mientras que al mismo tiempo grandes cantidades de iones positivos de potasio difunden hacia el exterior de la fibra. Estos dos efectos reducen el potencial intracelular hasta devolverlo a su nivel de reposo negativo y, por tanto, ponen fin al potencial de acción. Además, los canales de potasio permanecen abiertos durante algunas décimas de segundo más, manteniendo transitoriamente el movimiento de cargas positivas hacia el exterior de la célula, con el consiguiente exceso de negatividad en el interior de la fibra; este proceso se denomina hiperpolarización. El estado de hiperpolarización inicialmente desplaza el potencial de membrana «en reposo» hacia abajo hasta aproximadamente –55 o –60 mV al final del potencial de acción. Pero, ¿por qué este nuevo estado de hiperpolarización no se mantiene indefinidamente? El motivo es que en las décimas de segundo siguientes al final del potencial de acción se cierran cada vez más canales de potasio. Los iones sodio y calcio que fluyen hacia el interior una vez más compensan el flujo de salida de iones potasio, lo que lleva a que el potencial «de reposo» se desplace hacia arriba una vez más, alcanzando finalmente el nivel liminal de aproximadamente –40 mV. Después comienza de nuevo todo el proceso: autoexcitación para generar el potencial de acción, recuperación del potencial de acción, hiperpolarización después de que haya finalizado el potencial de acción, desplazamiento del potencial «de reposo» hasta el umbral y, finalmente, reexcitación para generar un nuevo ciclo. Este proceso continúa indefinidamente durante toda la vida de una persona.
LAS VÍAS INTERNODULARES E INTERAURICULARES TRANSMITEN IMPULSOS CARDÍACOS A TRAVÉS DE LAS AURÍCULAS
Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes. Por tanto, los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares. De esta manera, el potencial de acción se propaga por toda la, masa muscular auricular y, finalmente, llega hasta el nódulo AV. La velocidad de conducción en la mayor, parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s, en varias pequeñas bandas de fibras auriculares. Una de estas bandas, denominada banda interauricular anterior, atraviesa las paredes anteriores de las aurículas para dirigirse hacia la aurícula izquierda. Además, otras tres bandas pequeñas se incurvan a través de las paredes auriculares anterior, lateral y posterior, y terminan en el nódulo AV; y se denominan, respectivamente, vías internodulares anterior, media y posterior.
La causa de la velocidad de conducción más rápida de estas bandas es la presencia de fibras de conducción especializadas. Estas fibras son similares a las «fibras de Purkinje» de los ventrículos, que conducen incluso más rápidamente.
EL NÓDULO AURICULOVENTRICULAR RETRASA LA CONDUCCIÓN DEL IMPULSO DESDE LAS AURÍCULAS A LOS VENTRÍCULOS.
El sistema de conducción auricular está organizado de modo que el impulso cardíaco no viaja desde las aurículas hacia los ventrículos demasiado rápidamente; este retraso da tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular. El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes.
El nódulo AV está localizado en la pared posterolateral de la aurícula derecha, inmediatamente detrás de la válvula tricúspide, muestra en forma de diagrama las diferentes partes de este nódulo, más sus conexiones con las fibras de las vías internodulares auriculares entrantes y el haz AV de salida. Esta figura también muestra los intervalos temporales aproximados en fracciones de segundo entre el comienzo inicial del impulso cardíaco en el nódulo sinusal y su posterior aparición en el sistema del nódulo AV. Obsérvese que el impulso, después de viajar por las vías internodulares, llega al nódulo AV aproximadamente 0,03 s después de su origen en el nódulo sinusal. Después hay un retraso de otros 0,09 s en el propio nódulo AV antes de que el impulso entre en la porción penetrante del haz AV, a través del cual pasa hacia los ventrículos. Se produce un retraso final de otros 0,04 s principalmente en este haz AV penetrante, que está formado por múltiples fascículos pequeños que atraviesan el tejido fibroso que separa las aurículas de los ventrículos.
Así, el retraso total en el nódulo AV y en el sistema de AV es de aproximadamente 0,13 s. Este retraso, añadido al retraso inicial de la conducción de 0,03 s desde el nódulo sinusal hasta el nódulo AV, hace que haya un retraso total de 0,16 s antes de que la señal excitadora llegue finalmente al músculo ventricular que se está contrayendo.
Causa de la conducción lenta
La conducción lenta en las fibras transicionales, nodulares y penetrantes del haz AV está producida principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente. Por tanto, es fácil ver por qué se tarda en excitar células sucesivas.
TRANSMISIÓN RÁPIDA EN EL SISTEMA DE PURKINJE VENTRICULAR
Las fibras de Purkinje especiales se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Excepto en la porción inicial de estas fibras, donde penetran en la barrera fibrosa AV, tienen características funcionales bastante distintas a las de las fibras del nódulo AV. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad de 1,5 a 4 m/s, una velocidad aproximadamente seis veces mayor que la del músculo ventricular normal y 150 veces mayor que la de algunas de las fibras del nódulo AV. Esta velocidad permite una transmisión casi instantánea del impulso cardíaco por todo el resto del músculo ventricular.
Se piensa que la rápida transmisión de los potenciales de acción por las fibras de Purkinje está producida por un gran aumento del nivel de permeabilidad de las uniones en hendidura de los discos intercalados entre las células sucesivas que componen las fibras de Purkinje. Por tanto, los iones pasan fácilmente de una célula a la siguiente, aumentando de esta manera la velocidad de la transmisión. Las fibras de Purkinje también tienen muy pocas miofibrillas, lo que significa que se contraen poco o nada durante la transmisión de los impulsos.
Conducción unidireccional a través del haz AV
Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Esta característica impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo solo la contracción anterógrada desde las aurículas hacia los ventrículos.
Además, se debe recordar que en todas las localizaciones excepto en el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua. Esta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV. (En casos infrecuentes un puente muscular anormal penetra en la barrera fibrosa en otra localización distinta al haz AV. En estas condiciones el impulso cardíaco puede entrar en las aurículas desde los ventrículos y producir arritmias cardíacas graves.)
Distribución de las fibras de Purkinje en los ventrículos: las ramas izquierda y derecha del haz
Después de penetrar en el tejido fibroso que está entre el músculo auricular y ventricular, la porción distal del haz AV se dirigía hacia abajo en el interior del tabique interventricular a lo largo de 5 a 15 mm hacia la punta del corazón. Después el haz se divide en las ramas izquierda y derecha del haz, que están debajo del endocardio en los dos lados respectivos del tabique interventricular. Cada una de las ramas se dirige hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas. Estas ramas, a su vez, siguen un trayecto en dirección lateral alrededor de cada una de las cavidades ventriculares y hacia atrás, hacia la base del corazón. Los extremos de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas.
Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de solo 0,03 s. Por tanto, una vez que el impulso cardíaco ha entrado en el sistema de conducción ventricular de Purkinje, se propaga casi inmediatamente a toda la masa del músculo ventricular.
TRANSMISIÓN DEL IMPULSO CARDÍACO EN EL MÚSCULO VENTRICULAR
Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora solo de 0,3 a 0,5 m/s, una sexta parte de la velocidad de las fibras de Purkinje. El músculo cardíaco envuelve el corazón en una doble espiral, con tabiques fibrosos entre las capas en espiral; por tanto, el impulso cardíaco no viaja necesariamente directamente hacia el exterior, hacia la superficie del corazón, sino que se angula hacia la superficie a lo largo de las direcciones de las espirales. Debido a esta inclinación, la transmisión desde la superficie endocárdica a la superficie, epicárdica del ventrículo precisa hasta otros 0,03 s, aproximadamente igual al tiempo necesario para la, transmisión por toda la porción ventricular del sistema de Purkinje. Así, el tiempo total para la transmisión del impulso cardíaco desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular en el corazón normal es de aproximadamente 0,06 s
CONTROL DE LA EXCITACIÓN Y LA CONDUCCIÓN EN EL CORAZÓN
EL NÓDULO SINUSAL ES EL MARCAPASOS NORMAL DEL CORAZÓN
En el análisis que se ha realizado hasta ahora sobre la génesis y transmisión del impulso cardíaco por el corazón hemos señalado que el impulso normalmente se origina en el nódulo sinusal. En algunas situaciones anormales no ocurre así. Otras partes del corazón pueden presentar también una excitación rítmica intrínseca de la misma forma que lo hacen las fibras del nódulo sinusal; esta capacidad es particularmente cierto en el caso de las fibras del nódulo AV y de las fibras de Purkinje.
Las fibras del nódulo AV, cuando no son estimuladas por alguna fuente externa, descargan a una frecuencia rítmica intrínseca de 40 a 60 veces por minuto, y las fibras de Purkinje lo hacen a una frecuencia de entre 15 y 40 veces por minuto. Estas frecuencias son distintas a la frecuencia normal del nódulo sinusal, de 70 a 80 veces por minuto.
¿Por qué entonces es el nódulo sinusal, y no el nódulo AV ni las fibras de Purkinje, el que controla la ritmicidad del corazón? La respuesta procede del hecho de que la frecuencia de descarga del nódulo sinusal es considerablemente mayor que la frecuencia de descarga autoexcitadora natural de las fibras del nódulo AV y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Sin embargo, el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo AV o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Por tanto, el nuevo impulso procedente del nódulo sinusal descarga tanto las fibras del nódulo AV como las fibras de Purkinje antes de que se pueda producir autoexcitación en cualquiera de esas estructuras.
Así, el nódulo sinusal controla el latido del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón. Por tanto, el nódulo sinusal es casi siempre el marcapasos del corazón normal.
Marcapasos anormales: marcapasos «ectópico»
De manera ocasional alguna otra parte del corazón muestra una frecuencia de descarga rítmica que es más rápida que la del nódulo sinusal. Por ejemplo, a veces se produce este desarrollo en el nódulo AV o en las fibras de Purkinje cuando una de estas estructuras se altera. En ambos casos el marcapasos del corazón se desplaza desde el nódulo sinusal hasta el nódulo AV o las fibras de Purkinje excitadas. En casos menos frecuentes todavía, algún punto del músculo auricular o ventricular presenta una excitabilidad excesiva y se convierte en el marcapasos.
Un marcapasos que está situado en una localización distinta al nódulo sinusal se denomina marcapasos «ectópico». Un marcapasos ectópico da lugar a una secuencia anormal de contracción de las diferentes partes del corazón y puede producir una debilidad significativa del bombeo cardíaco.
Otra causa de desplazamiento del marcapasos es el bloqueo de la transmisión del impulso cardíaco desde el nódulo sinusal a las demás partes del corazón. El nuevo marcapasos se produce en este caso con más frecuencia en el nódulo AV o en la porción penetrante del haz AV en su trayecto hacia los ventrículos.
Cuando se produce un bloqueo AV, es decir, cuando el impulso cardíaco no puede pasar desde las aurículas hacia los ventrículos a través del sistema del nódulo AV y del haz, las aurículas siguen latiendo a la frecuencia normal del ritmo del nódulo sinusal, mientras que habitualmente aparece un nuevo marcapasos en el sistema de Purkinje de los ventrículos que activa el músculo ventricular a una frecuencia de entre 15 y 40 latidos/min. Después de un bloqueo súbito del haz AV el sistema de Purkinje no comienza a emitir sus impulsos rítmicos intrínsecos hasta 5 a 20 s después porque, antes del bloqueo, las fibras de Purkinje habían estado «sobreexcitadas» por los rápidos impulsos sinusales y, en consecuencia, están en un estado suprimido. Durante estos 5 a 20 s los ventrículos dejan de bombear sangre y la persona se desvanece después de los primeros 4 a 5 s debido a la ausencia de flujo sanguíneo cerebral. Este retraso de la recuperación del corazón se denomina síndrome de Stokes-Adams. Si el período de retraso es demasiado largo, se puede producir la muerte.
LOS NERVIOS SIMPÁTICOS Y PARASIMPÁTICOS CONTROLAN EL RITMO CARDÍACO Y LA CONDUCCIÓN DE IMPULSOS POR LOS NERVIOS CARDÍACOS
El corazón está inervado por nervios simpáticos y parasimpáticos. Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV, en mucho menor grado al músculo de las dos aurículas y apenas directamente al músculo ventricular. Por el contrario, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas.
La estimulación parasimpática (vagal) ralentiza el ritmo y la conducción cardíacos
La estimulación de los nervios parasimpáticos que llegan al corazón (los vagos) hace que se libere la hormona acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón. Primero, reduce la frecuencia del ritmo del nódulo sinusal, y segundo, reduce la excitabilidad de las fibras de la unión AV entre la musculatura auricular y el nódulo AV, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos.
Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal. Además, la estimulación intensa de los nervios vagos puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo AV. En cualquiera de los casos, las señales excitadoras rítmicas ya no se transmiten hacia los ventrículos. Los ventrículos pueden dejar de latir durante 5 a 20 s, pero después alguna área pequeña de las fibras de Purkinje, habitualmente en la porción del tabique interventricular del haz AV, presenta un ritmo propio y genera la contracción ventricular a una frecuencia de 15 a 40 latidos/min. Este fenómeno se denomina escape ventricular.
Mecanismo de los efectos vagales
La acetilcolina que se libera en las terminaciones nerviosas vagales aumenta mucho la permeabilidad de las membranas de las fibras a los iones potasio, lo que permite la salida rápida de potasio desde las fibras del sistema de conducción. Este proceso da lugar a un aumento de la negatividad en el interior de las fibras, un efecto que se denomina hiperpolarización, que hace que este tejido excitable sea mucho menos excitable.
En el nódulo sinusal, el estado de hiperpolarización hace el potencial de membrana «en reposo» de las fibras del nódulo sinusal mucho más negativo de lo habitual, es decir, de –65 a –75 mV en lugar del nivel normal de –55 a –60 mV. Por tanto, el aumento inicial del potencial de membrana del nódulo sinusal que produce la corriente de entrada de sodio y de calcio tarda mucho más en alcanzar el potencial liminal para la excitación. Este requisito retrasa mucho la frecuencia de ritmicidad de estas fibras nodulares. Si la estimulación vagal es lo suficientemente intensa es posible detener totalmente la autoexcitación rítmica de este nódulo.
En el nódulo AV, el estado de hiperpolarización producido por la estimulación vagal hace que sea difícil que las pequeñas fibras auriculares que entran en el nódulo generen una corriente de una intensidad suficiente como para excitar las fibras nodulares. Por tanto, el factor de seguridad para la transmisión del impulso cardíaco a través de las fibras de transición hacia las fibras del nódulo AV disminuye. Una reducción moderada simplemente retrasa la conducción del impulso, aunque una disminución grande bloquea totalmente la conducción.
La estimulación simpática aumenta el ritmo y la conducción del corazón
La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal. Primero, aumenta la frecuencia de descarga del nódulo sinusal. Segundo, aumenta la velocidad de conducción, así como el nivel de excitabilidad de todas las porciones del corazón. Tercero, aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular.
En breve, la estimulación simpática aumenta la actividad global del corazón. La estimulación máxima casi puede triplicar la frecuencia del latido cardíaco y puede aumentar la fuerza de la contracción del corazón hasta dos veces.
Mecanismo del efecto simpático
La estimulación de los nervios simpáticos libera la hormona noradrenalina en las terminaciones nerviosas simpáticas. La noradrenalina estimula, a su vez, los receptores β1-adrenérgicos, que median en los efectos sobre la frecuencia cardíaca. No está del todo claro el mecanismo preciso mediante el que la estimulación β1-adrenérgica actúa sobre las fibras del músculo cardíaco, aunque se piensa que aumenta la permeabilidad de la membrana de las fibras a los iones sodio y calcio. En el nódulo sinusal, un aumento de la permeabilidad a sodio-calcio genera un potencial en reposo más positivo y también produce un aumento de la velocidad del ascenso del potencial de membrana diastólico hacia el nivel liminal para la autoexcitación, acelerando de esta forma la autoexcitación y, por tanto, aumentando la frecuencia cardíaca.
En el nódulo AV y en los haces AV, el aumento de la permeabilidad a sodio-calcio hace que sea más fácil que el potencial de acción excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo de esta manera el tiempo de conducción desde las aurículas hasta los ventrículos. El aumento de la permeabilidad a los iones calcio es responsable al menos en parte del aumento de la fuerza contráctil del músculo cardíaco bajo la influencia de la estimulación simpática, porque los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas.
El corazón tiene dos bombas: un corazón derecho que se encarga de recibir sangre de los órganos periféricos y llevarlos hacia los pulmones. Un corazón izquierdo que recibe la sangre oxigenada de los pulmones y los bombea de nuevo hacia los órganos periféricos. Cada una de estas se forma por una aurícula y un ventrículo; la primera es la encargada de llenar los ventrículos de sangre mientras que el segundo tiene la función de expulsarla hacia la circulación.
Características distintivas del músculo cardíaco en relación con el músculo esquelético.
Los músculos cardiaco y esquelético son estriados y contienen filamentos actina y miosina.
El cardiaco posee discos intercalados entre las células musculares, las cuales tienen un potencial de acción bajo que permite que este viaje por todas las células musculares.
El musculo cardiaco es un sincitio y su haz auriculoventricular (AV) conduce lentamente los impulsos desde las aurículas a los ventrículos.
El potencial del músculo cardiaco en reposo es de -85 a -95 mV y su potencial es de 105 mV.
La entrada lenta de iones sodio y calcio en las células musculares cardiacas es una de las causas de la meseta del potencial de acción, ya que contiene canales lentos de calcio exclusivo o canales de sodio-calcio.
Los iones entran dichos canales y provocan la contracción del músculo.
Otra causa de la meseta del potencial de acción es el descenso de la permeabilidad de las células del músculo cardíaco a los iones de potasio, es decir, cuando los canales lentos de sodio-calcio se cierran después de 0,2 a 0,3 s, aumenta la permeabilidad de la membrana a los iones potasio y por lo tanto el potencial de membrana vuelve a su nivel de reposo.
Otra característica es que la difusión del calcio en las miofibrillas promueve la contracción muscular.
El potencial se propaga por los túbulos transversos (T) haciendo que los túbulos longitudinales liberen iones calcio al retículo sarcoplasmatico, lo cual favorecen el deslizamiento de los filamentos de actina y miosina lo que da lugar a la contracción muscular.
Ciclo cardiáco.
El fenómeno se inicia desde el comienzo de un latido cardíaco hasta el comienzo del siguiente a lo que se denomina ciclo cardíaco.
Cada ciclo es inicio por la generación espontánea de un potencial de acción en el nódulo sinusal en la aurícula derecha.
El potencial se desplaza a través de ambas aurículas, mientras que el nodo y el haz AV hacia los ventrículos, existe un retraso de 0,1 s lo que permite que las aurículas se contraigan antes que los ventrículos.
Los ventrículos se llenan de sangre durante la diástole y se contrae durante la sístole
La propagación del potencial de acción se da a cada latido, para su registro está el electrocardiograma el cual registra el voltaje generado por el corazón. La onda P está producida por la despolarización de las aurículas, que causa la contracción de esta. Las ondas QRS aparecen en consecuencia de la despolarización ventricular aproximadamente 0,16s después del inicio de la onda P y marca el comienzo de contracción ventricular. La onda T ventricular se debe a repolarización del ventrículo.
Las aurículas funcionan como bombas de cebado para los ventrículos. El 75% del llenado ventricular se produce durante la diástole antes de la contracción de las aurículas, que causa el 25% del llenado ventricular. Algunas de las ondas ventriculares son las siguientes: la onda a que se debe a la contracción auricular, la onda c que se produce durante la contracción ventricular, la onda v que se debe al flujo lento de sangre hacia las aurículas por el retorno venoso.
Los ventrículos se llenan con sangre durante la diástole, el comienzo de esta es el periodo de relajación isovolumétrica, provocando relajación ventricular y las válvulas AV se abren, la presión más alta hace que las aurículas empujen la sangre hacia los ventrículos, luego ocurre el llenado ventricular y por último la contracción auricular que contribuye al 25% del llenado.
Durante el comienzo de la sístole se cierran la contracción ventricular, las válvulas AV se cierran y la presión empieza subir, sin embargo las válvulas de la aortica y pulmonar se abren con la presión en el ventrículo izquierdo de 80mmHg y el ventrículo derecho con presión de 8mmHg, a este periodo se le llama periodo de inyección. En la última fase de la sístole las presiones bajan a comparación de las antes mencionadas provocando que as válvulas semilunares se cierren.
El trabajo sistólico de los ventrículos es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco. El trabajo volumen-presión el corazón es el trabajo realizado para aumentar la presión en la sangre.
El ciclo cardiaco presenta ciertas fases las cuales son las siguientes: la fase 1 es de llenado durante el cual el volumen ventricular izquierdo aumenta desde un volumen telesistólico a uno telediastólico. La fase dos consiste en el periodo de refracción isovolumetrica, durante la cual el volumen del ventrículo se mantiene pero la presión interventricular aumenta a un nivel de la presión diastólica aortica. La fase tres es el periodo de inyección durante la cual la presión sistólica aumenta de nuevo debido a una contracción ventricular adicional. La fase cuatro es el periodo de relajación isovolumetrica.
El consumo de oxígeno en el corazón dependerá del trabajo cardiaco que es el proceso de volumen-presión, este consumo de oxigeno es proporcional a la tensión del corazón multiplicada por el tiempo en que se mide la tensión.
El mecanismo de Frank-Stirling
regula intrínsecamente la capacidad de bomba cardiaca, es decir cuando el retorno venoso aumenta, el musculo se estira más y bombea con una fuerza de contracción mayor, en otro modo esta ley dice que dentro de los límites fisiológicos el corazón bombea toda la sangre que le llega sin dejar que se acumule el exceso de sangre en las venas.
El sistema nervioso autónomo afecta a la función de bomba cardiaca, cada estimulación intensa la frecuencia cardiaca de un adulto puede aumentar de una unidad de reposo de 72 latidos por minuto hasta 180-200 por min. La contractibilidad cardiaca depende de varios factores, como las contracciones de electrolitos extracelulares, algunos ejemplos para que se de este suceso es exceso de calcio que hace que el corazón provoque contracciones espásticas.

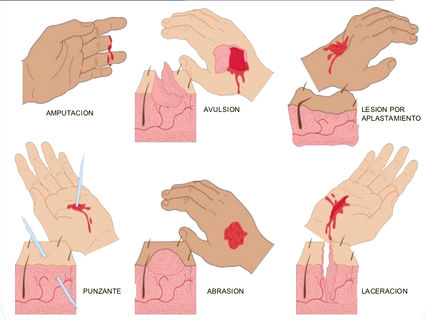
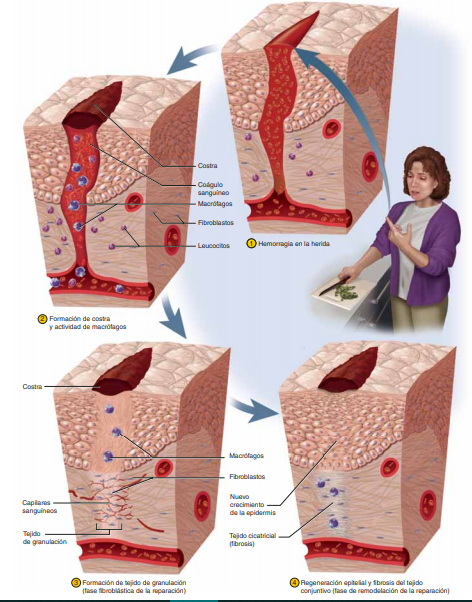
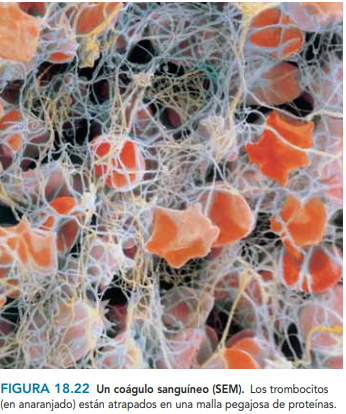
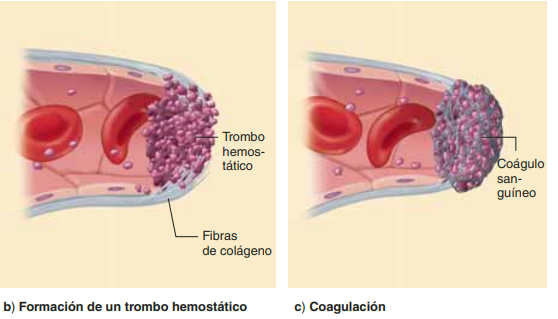
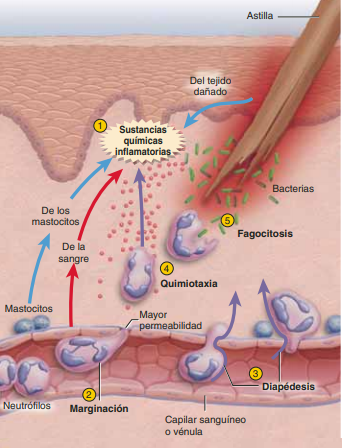
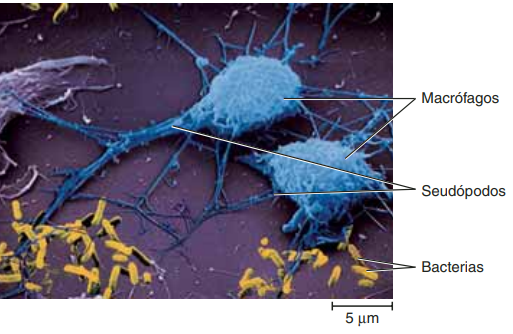





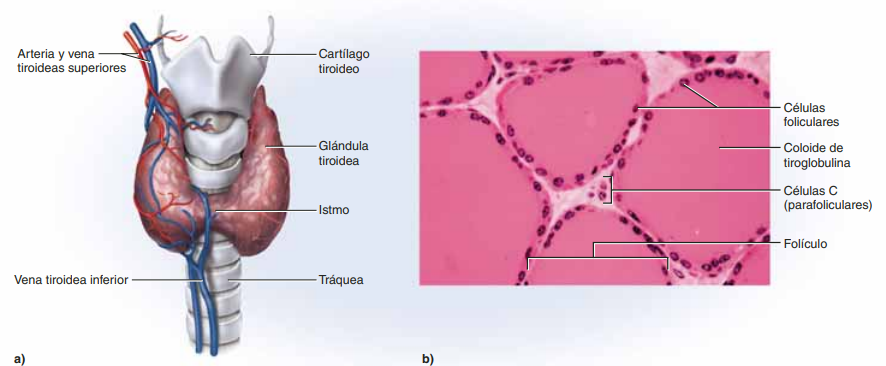
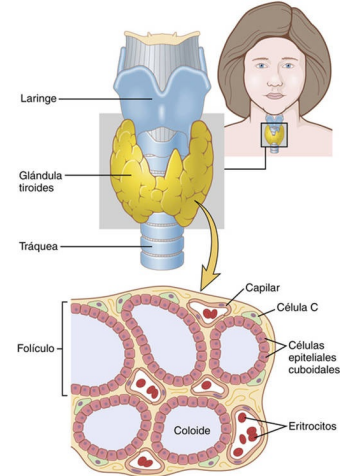
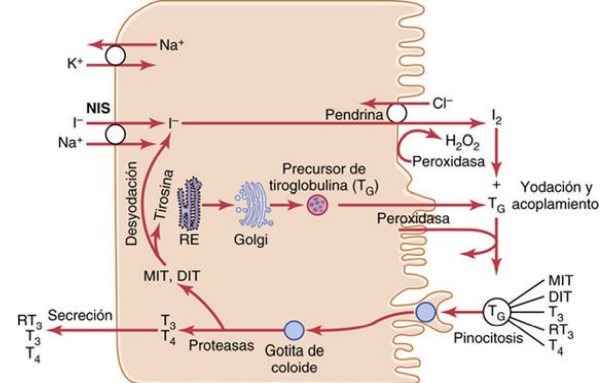
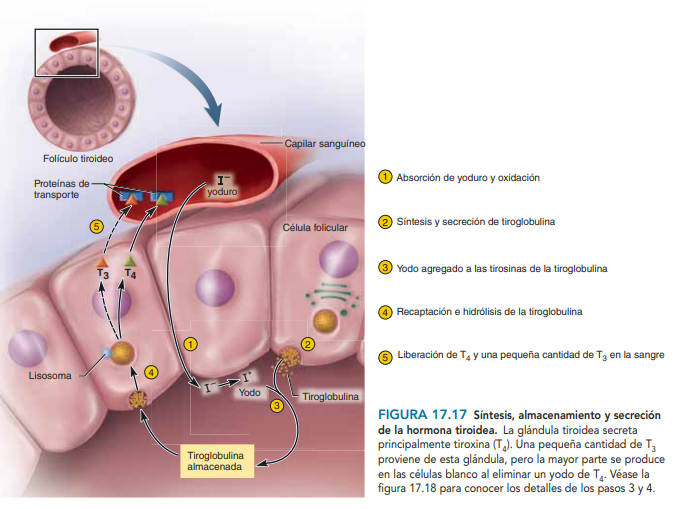
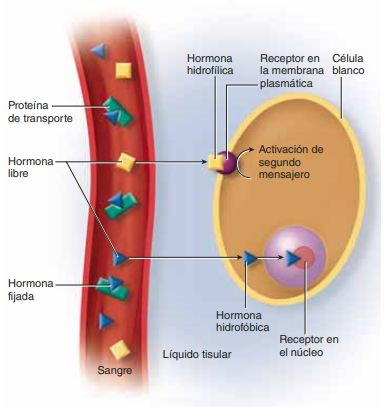
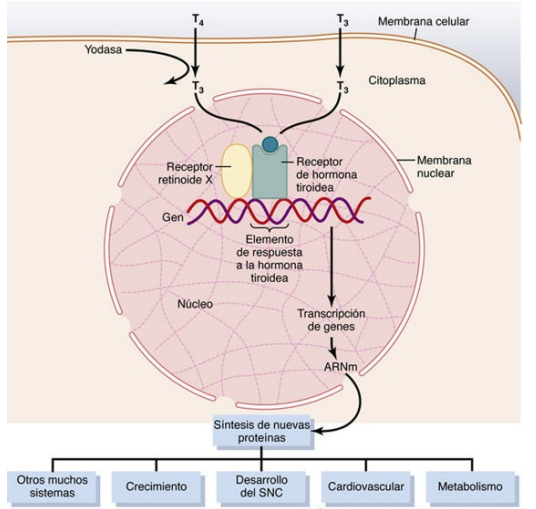



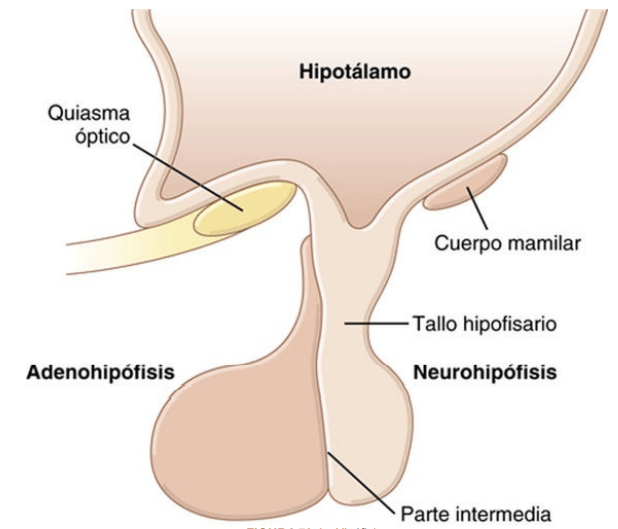
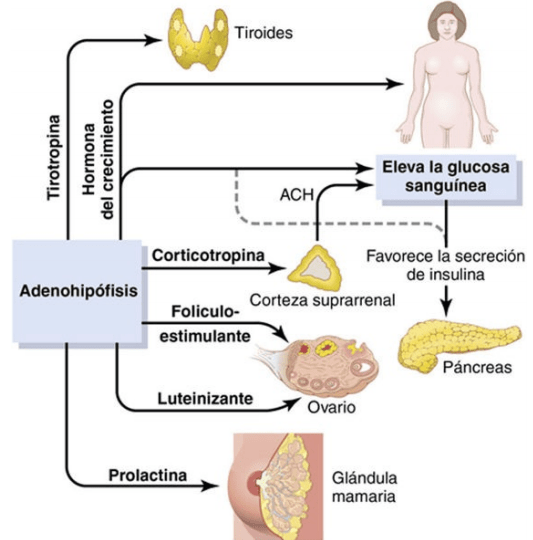
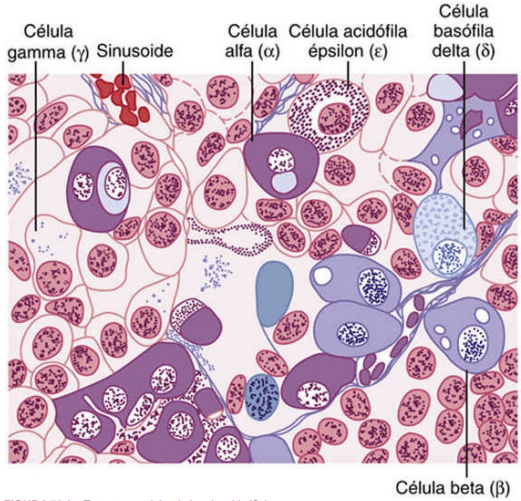
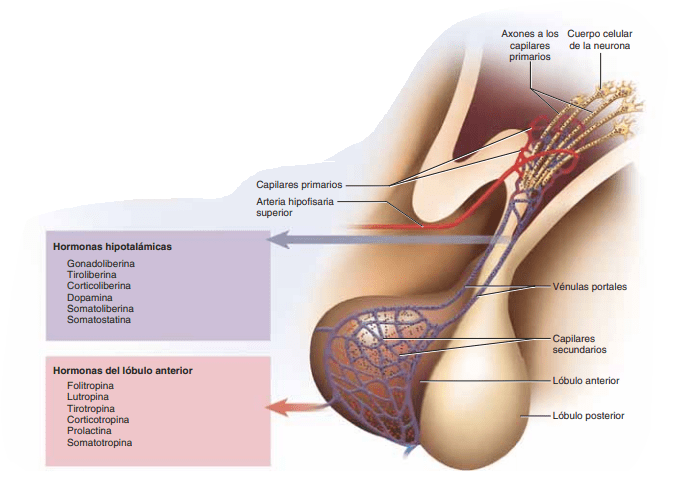
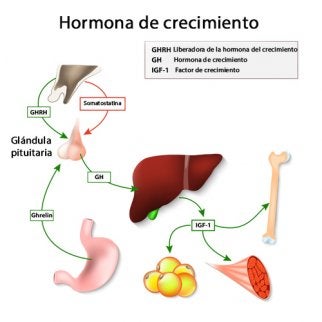
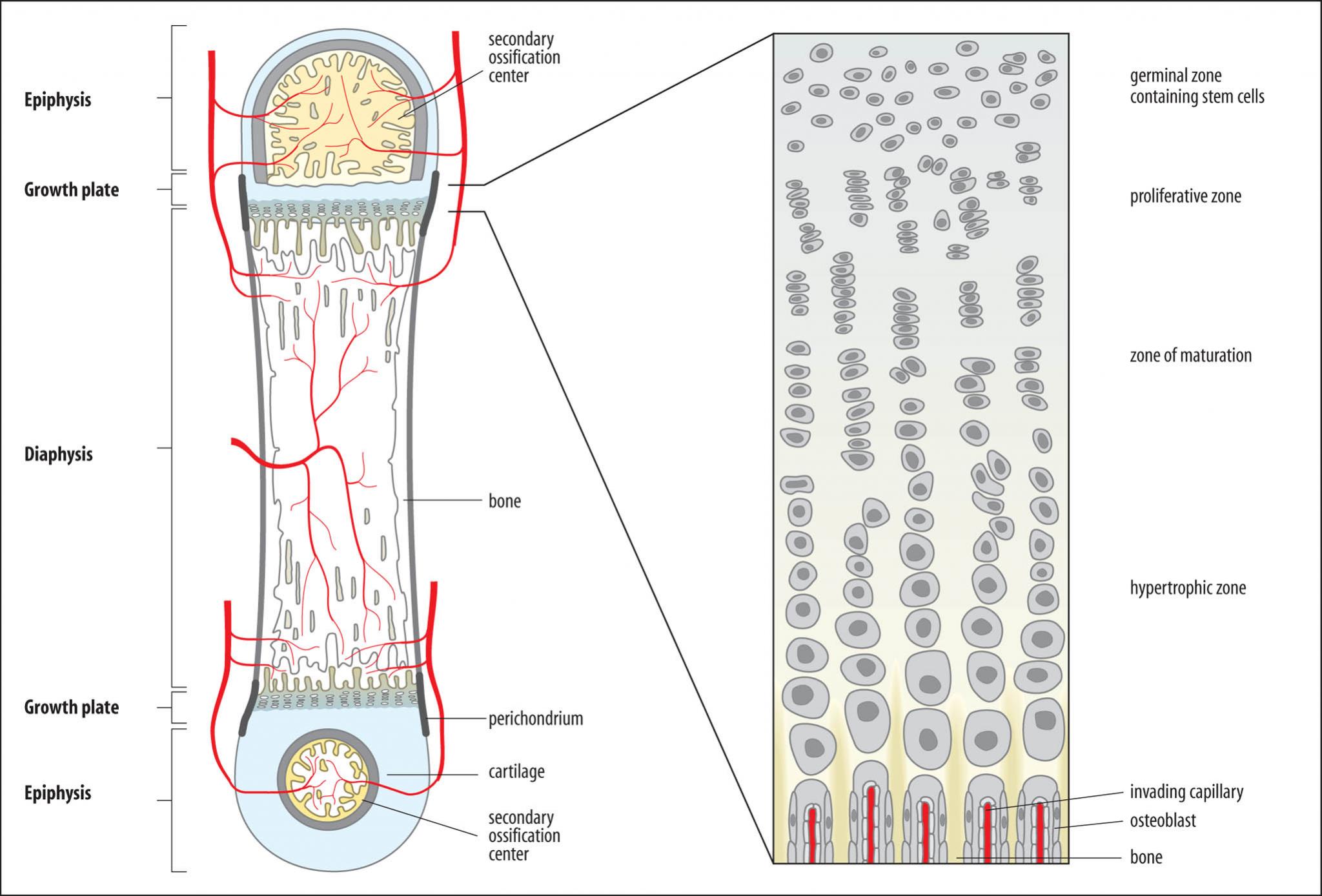



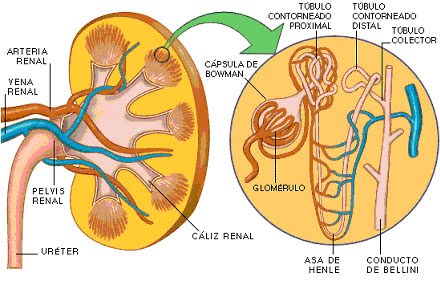
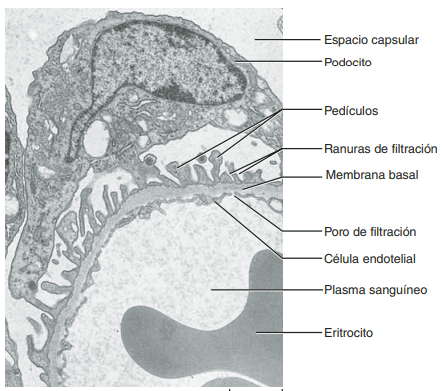
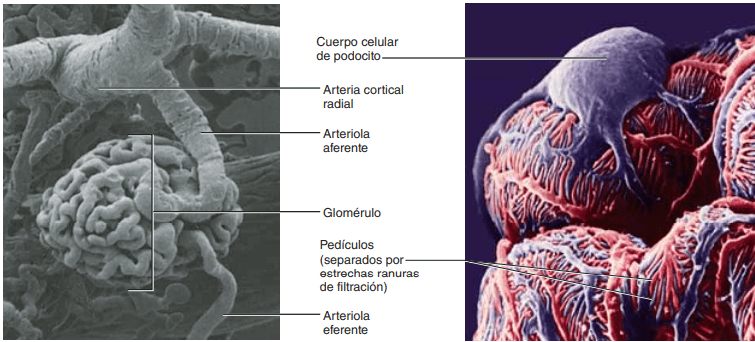
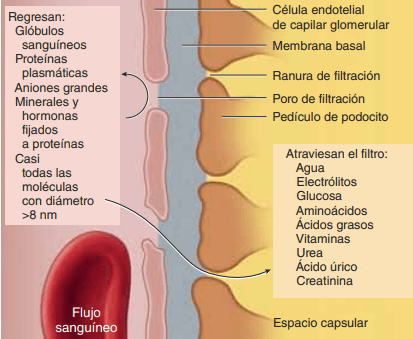
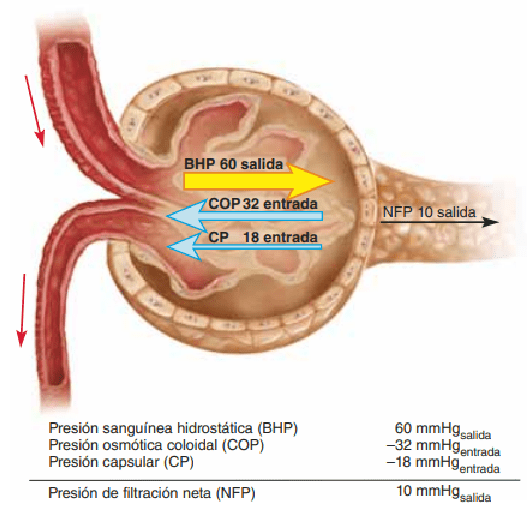
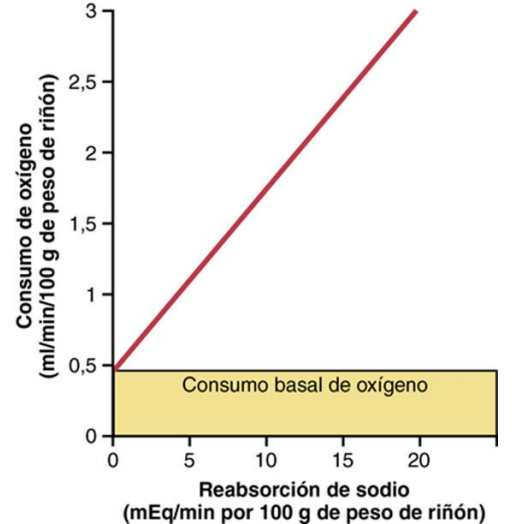
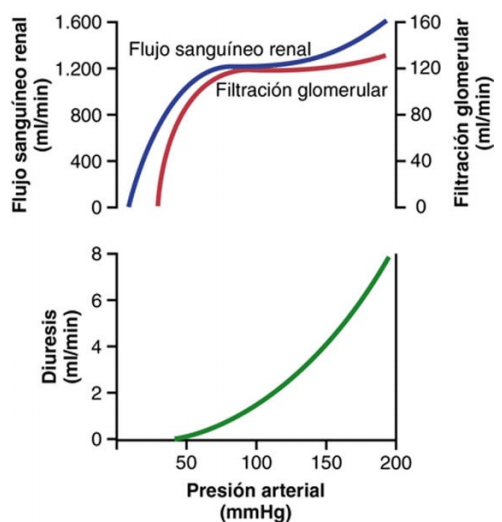
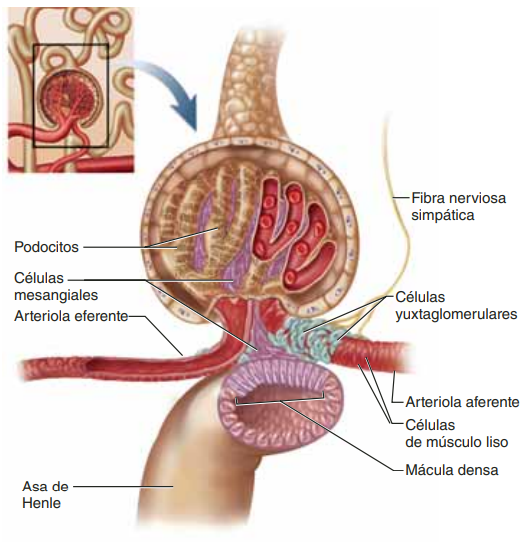
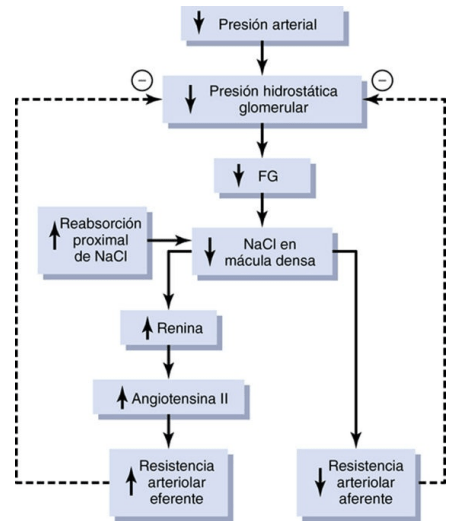
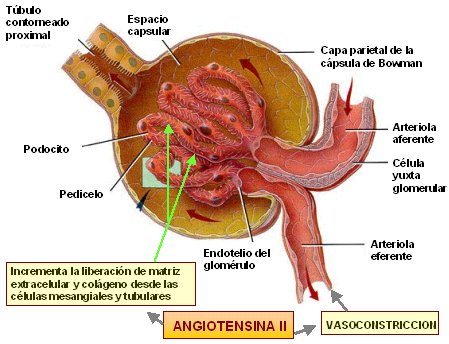

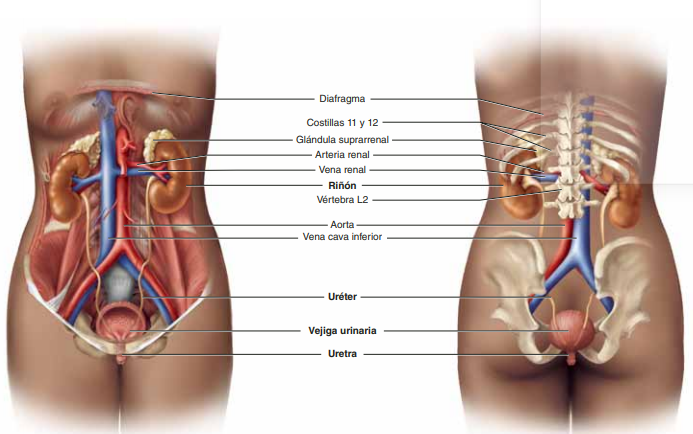
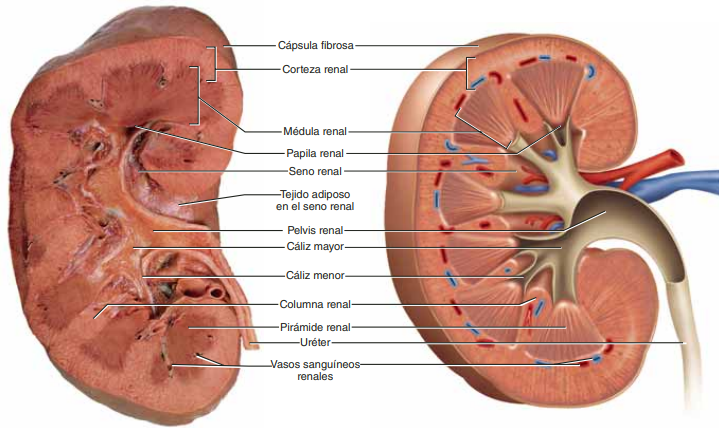
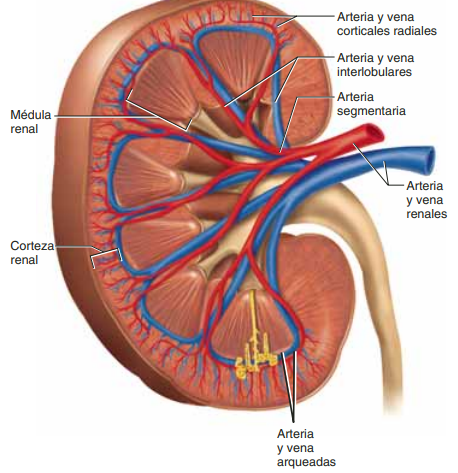
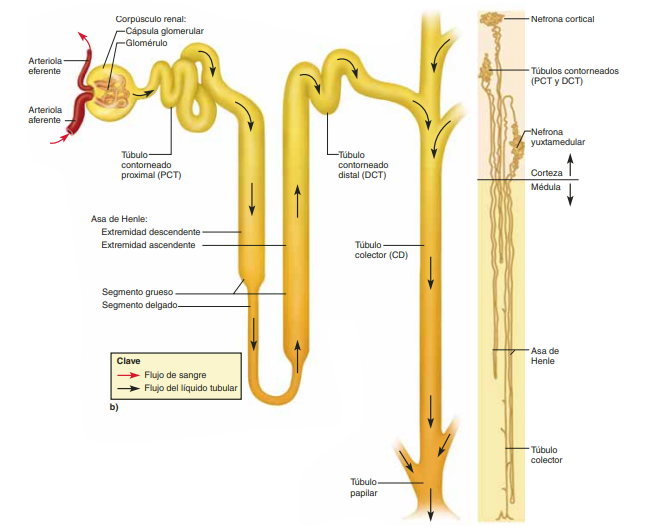
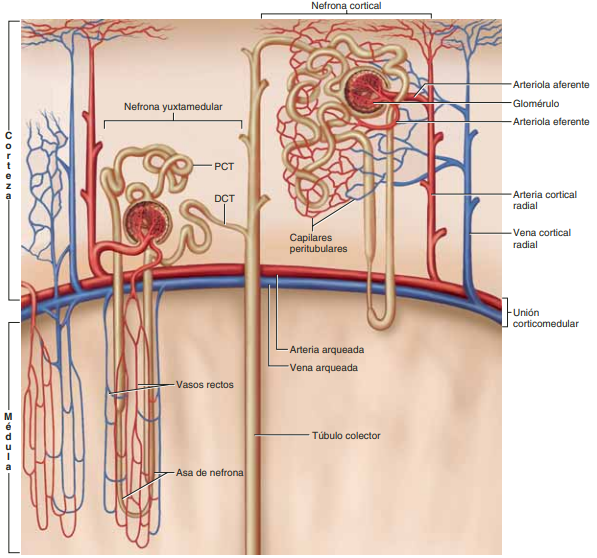
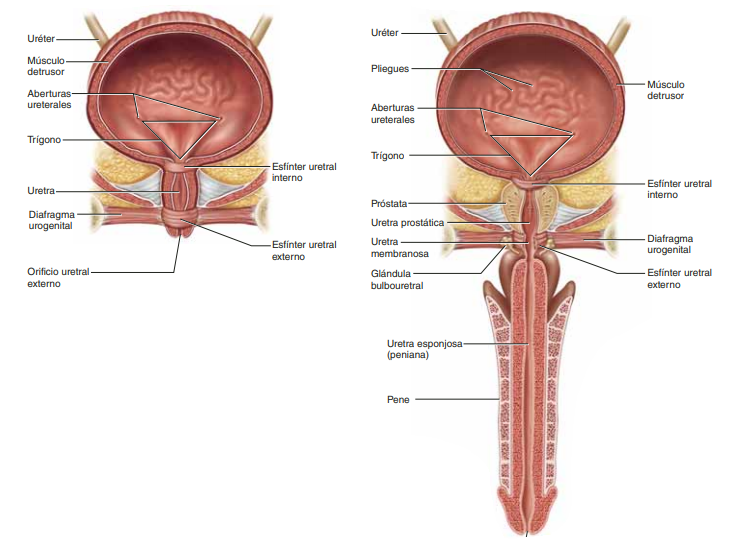

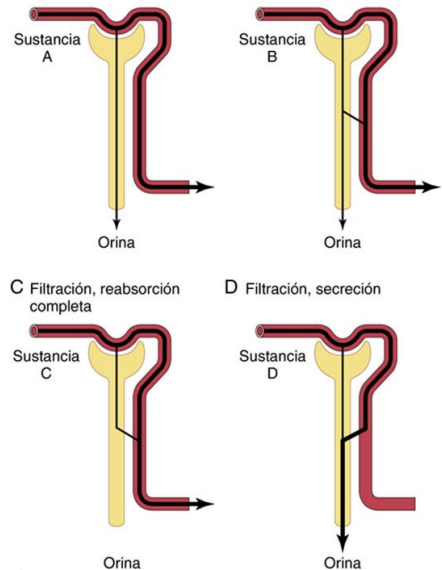


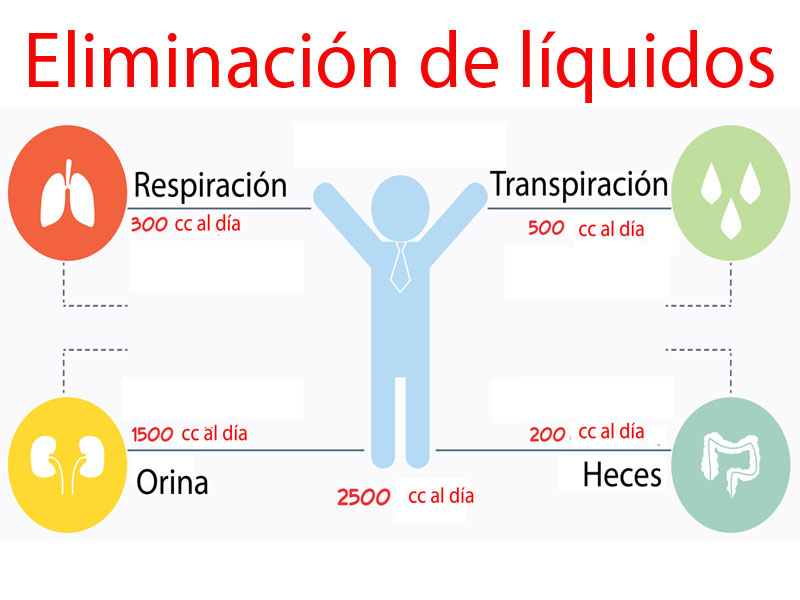

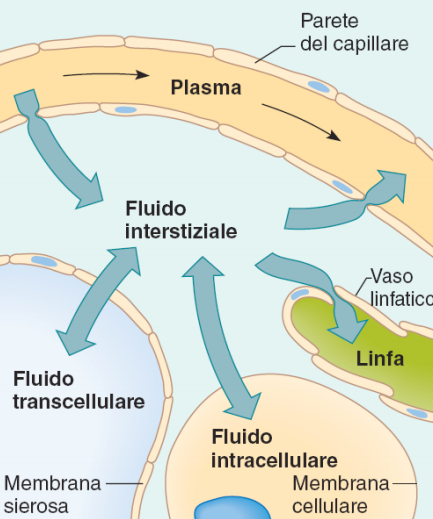
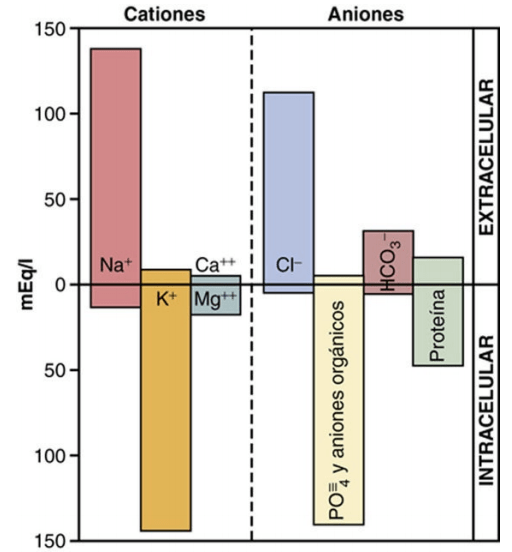

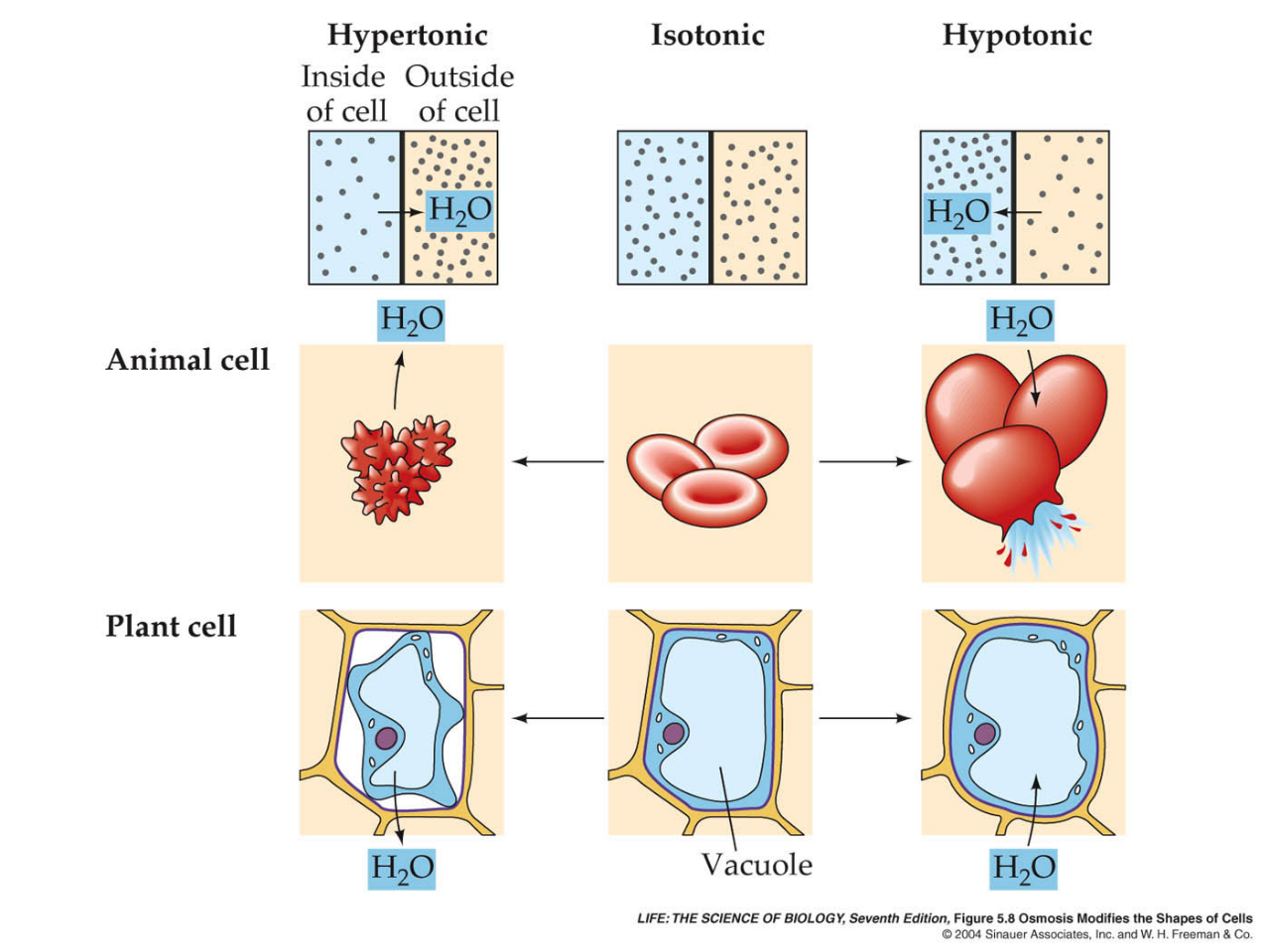






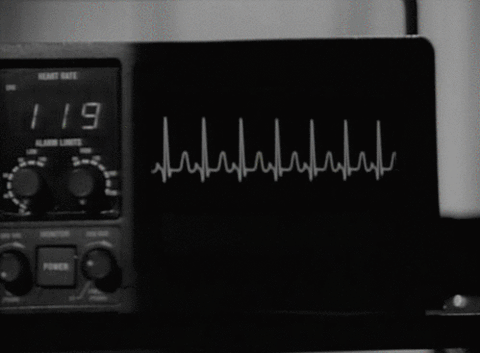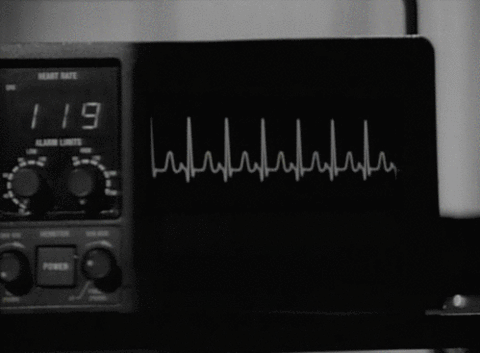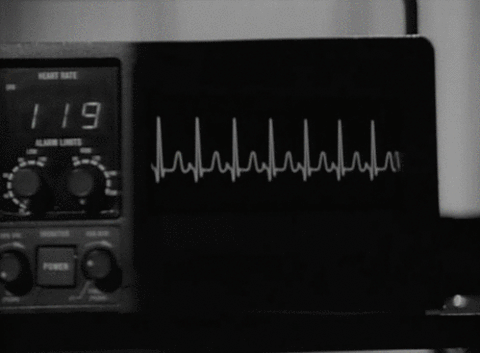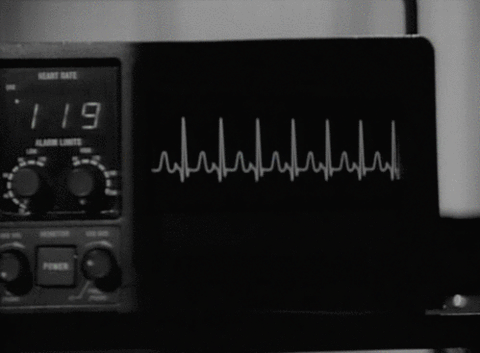



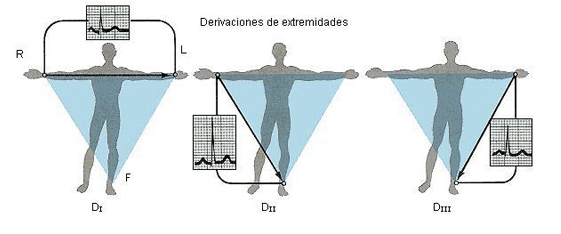

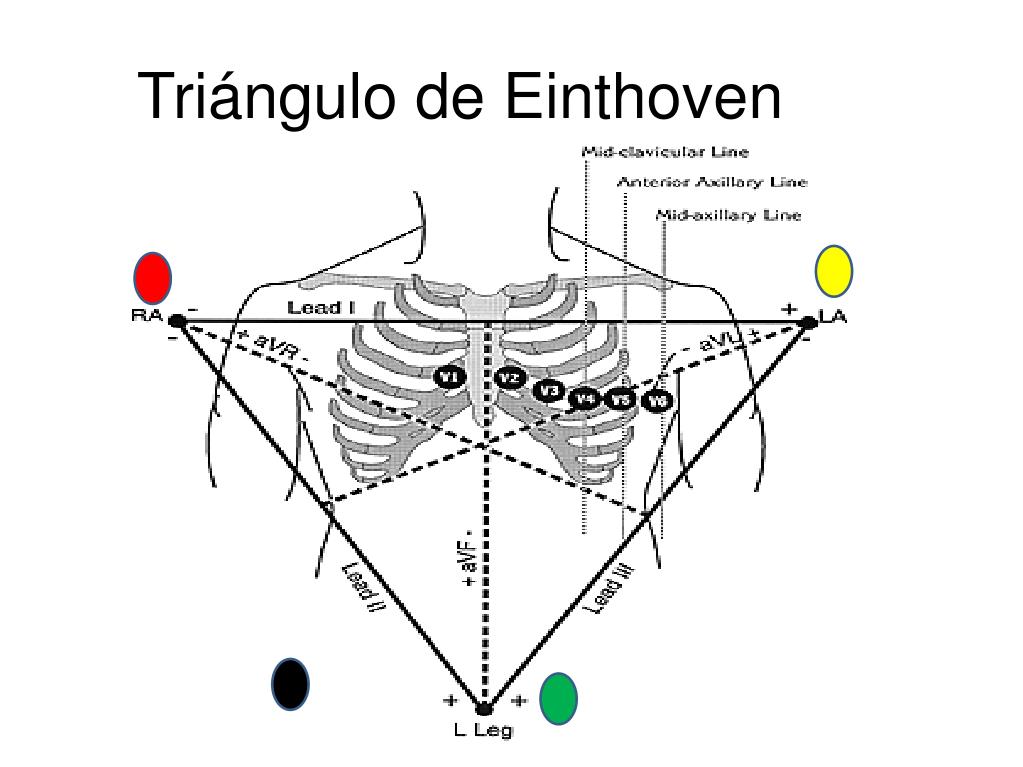
















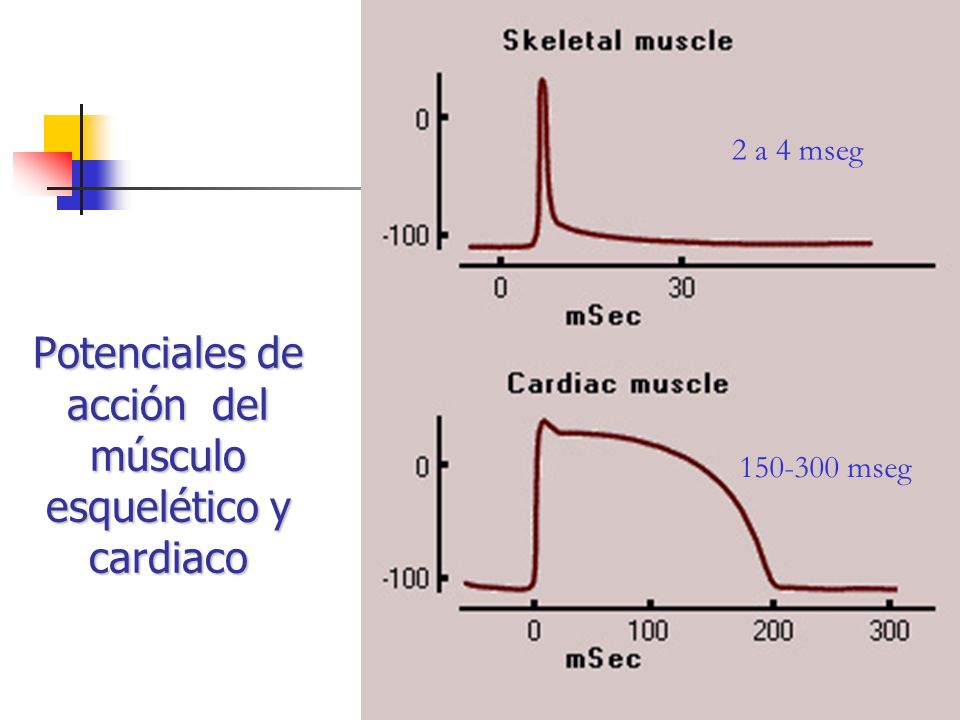

:max_bytes(150000):strip_icc()/Coraznconvasoscoronariosilustracin-5b16e4113de423003779afca.jpg)