La sensibilidad somática es el mecanismo nervioso que recopila la información sensitiva de todo el cuerpo
Clasificación de las sensibilidades somáticas
Las sensibilidades somáticas pueden clasificarse en tres tipos fisiológicos:
1) las sensibilidades somáticas mecanorreceptoras, formadas por las sensaciones táctiles y posicionales cuyo estímulo depende del desplazamiento mecánico de algún tejido del organismo.
2) las sensibilidades termorreceptoras, que detectan el calor y el frío.
3) la sensibilidad al dolor, que se activa con factores que dañan los tejidos.
Otras clasificaciones de las sensibilidades somáticas:
La sensibilidad exterorreceptora es la que procede de la superficie del cuerpo. La sensibilidad propioceptiva es la que tiene que ver con el estado físico del cuerpo, como las sensaciones posicionales, las tendinosas y musculares,La sensibilidad visceral es la que deriva de las vísceras del cuerpo;La sensibilidad profunda es la que viene de los tejidos profundos, como las fascias, los músculos y los huesos. Esta sensibilidad comprende básicamente la presión «profunda», el dolor y la vibración
Detección y transmisión de las sensaciones táctiles
Interrelaciones entre las sensaciones táctiles de contacto, presión y vibración Aunque el tacto, la presión y la vibración suelen clasificarse como sensaciones independientes, todas se detectan mediante los mismos tipos de receptores. Existen tres diferencias principales entre ellas:
1) la sensación de tacto en general deriva de la estimulación de los receptores táctiles situados en la piel o en los tejidos inmediatamente por debajo de ella;
2) la sensación de presión suele obedecer a la deformación de los tejidos profundos.
3) la sensación de vibración resulta de la repetición de señales sensitivas con rapidez, pero recurre a algunos tipos de receptores que también emplean las de tacto y de presión.
Vías sensitivas para la transmisión de señales somáticas en el sistema nervioso central
Casi toda la información sensitiva procedente de los segmentos somáticos corporales penetra en la médula espinal a través de las raíces dorsales de los nervios raquídeos. Sin embargo, desde su punto de entrada estas señales son transmitidas por la médula y más tarde por el encéfalo a través de una de las dos vías sensitivas alternativas siguientes: 1) el sistema de la columna dorsal-lemnisco medial, o 2) el sistema anterolateral.
El sistema de la columna dorsal-lemnisco medial:
como su nombre da a entender, transporta señales en sentido ascendente básicamente por las columnas dorsales de la médula hacia el bulbo raquídeo en el encéfalo. A continuación, después de hacer sinapsis y cruzar al lado opuesto a este nivel, siguen subiendo a través del tronco del encéfalo hasta el tálamo dentro del lemnisco medial.
El sistema anterolateral
nada más entrar en la médula espinal procedentes de las raíces dorsales de los nervios raquídeos, hacen sinapsis en las astas dorsales de la sustancia gris medular, después cruzan al lado opuesto y ascienden a través de sus columnas blancas anterior y lateral.
Anatomía del sistema de la columna dorsal-lemnisco medial
las fibras nerviosas que penetran en las columnas dorsales siguen su trayecto sin interrupción hasta la zona dorsal del bulbo raquídeo, donde hacen sinapsis en los núcleos de la columna dorsal (los núcleos cuneiforme y grácil). Desde aquí, las neuronas de segundo orden se decusan de inmediato hacia el lado opuesto del tronco del encéfalo y continúan ascendiendo a través de los lemniscos mediales hasta el tálamo. En su recorrido por el tronco del encéfalo, otras fibras nuevas procedentes de los núcleos sensitivos del nervio trigémino se incorporan a cada lemnisco medial; estas fibras desempeñan las mismas funciones sensitivas para la cabeza que las fibras de la columna dorsal para el cuerpo.
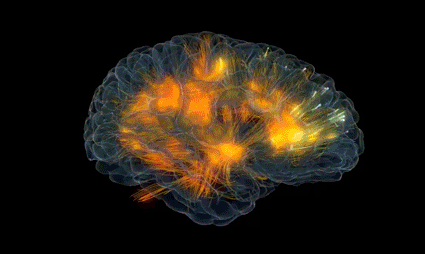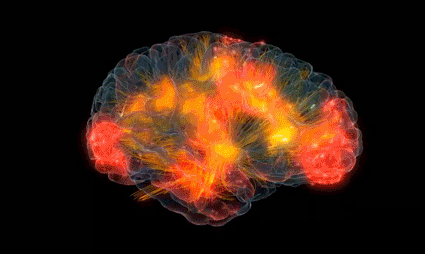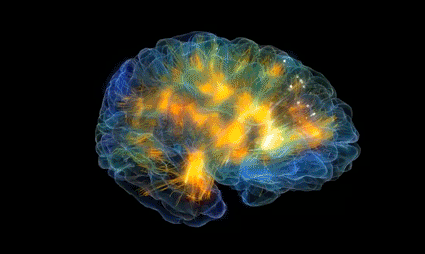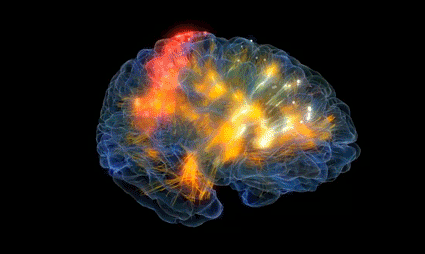
En el tálamo, las fibras del lemnisco medial terminan en la zona talámica de relevo sensitivo, llamada complejo ventrobasal. Desde este punto, las fibras nerviosas de tercer orden proyectan, según se muestra en la figura 48-4, sobre todo hacia la circunvolución poscentral de la corteza cerebral, que recibe el nombre de área sensitiva somática I
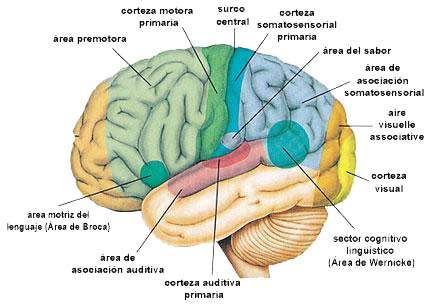
Corteza somatosensitiva
es un mapa de la corteza cerebral humana, que manifiesta su división en unas 50 zonas distintas llamadas áreas de Brodmann según su diferente estructura histológica
la gran cisura central (también llamada surco central) que se extiende en sentido horizontal cruzando el cerebro. En general, las señales sensitivas pertenecientes a cualquier modalidad de sensación terminan en la corteza cerebral inmediatamente por detrás de la cisura central. También a grandes rasgos, la mitad anterior del lóbulo parietal se ocupa casi por completo de la recepción e interpretación de las señales somatosensitivas, pero la mitad posterior aporta unos niveles aún más altos de interpretación. Las señales visuales acaban en el lóbulo occipital, y las señales auditivas terminan en el lóbulo temporal.
la porción de la corteza cerebral que queda delante de la cisura central y constituye la mitad posterior del lóbulo frontal se llama corteza motora y está dedicada casi en su integridad a controlar las contracciones musculares y los movimientos del cuerpo.
Orientación espacial de las señales procedentes de distintas partes del cuerpo en el área somatosensitiva I
El área somatosensitiva I se halla inmediatamente detrás de la cisura central, situada en la circunvolución poscentral de la corteza cerebral humana (corresponde a las áreas de Brodmann 3, 1 y 2).
Algunas regiones están representadas por grandes zonas en la corteza somática (la mayor corresponde a los labios, seguida por la cara y el pulgar), mientras que el tronco y la parte inferior del cuerpo están representados por una zona relativamente pequeña. Los tamaños de estos territorios son directamente proporcionales al número de receptores sensitivos especializados en cada zona periférica respectiva del organismo.
Capas de la corteza somatosensitiva y su función
La corteza cerebral contiene seis capas de neuronas, comenzando por la capa I próxima a la superficie cerebral y siguiendo cada vez por zonas más profundas hasta la capa VI
las neuronas de cada capa ejecutan funciones diferentes a las de las otras. Algunas de ellas son las siguientes
La señal sensitiva entrante excita en primer lugar la capa neuronal IV; a continuación, se propaga hacia la superficie de la corteza y también hacia otras capas más profundas.
. Las capas I y II reciben señales de entrada difusas inespecíficas procedentes de los centros inferiores del encéfalo, que facilitan regiones corticales específicas
Las neuronas de las capas II y III envían axones hacia las porciones emparentadas entre sí de la corteza cerebral en el lado opuesto del cerebro a través del cuerpo calloso.
Las neuronas de las capas V y VI mandan axones hacia las partes más profundas del sistema nervioso
Campos segmentarios de la sensación: dermatomas
Cada nervio raquídeo se encarga de un «campo segmentario» de la piel denominado dermatoma
Es posible emplear este recurso para determinar el nivel de la médula espinal en el que se ha producido una lesión medular cuando quedan alteradas las sensaciones periféricas por la lesión.
El dolor es un mecanismo que protege el organismo.
Aparece cada vez que se lesiona cualquier tejido y hace que el sujeto reaccione eliminando el estímulo doloroso.
CLASIFICACION DEL DOLOR.
El dolor rápido también se describe con otros muchos nombres alternativos, como dolor intenso, dolor punzante, dolor agudo y dolor eléctrico. Este tipo de dolor se siente cuando se clava una aguja en la piel, cuando se corta con un cuchillo o cuando sufre una quemadura intensa. También se percibe cuando la piel se ve sometida a una sacudida eléctrica
El dolor lento también se designa con muchos nombres, entre ellos dolor lento urente, dolor sordo, dolor pulsátil, dolor nauseoso y dolor crónico. Este tipo de dolor suele ir asociado a una destrucción tisular. El dolor lento tiene la capacidad de propiciar un sufrimiento casi insoportable y prolongado. Puede darse en la piel y casi también en cualquier tejido u órgano profundo.
RECEPTORES DEL DOLOR Y ESTIMULOS DOLOROSOS.
Son terminaciones nerviosas libres en la piel, tejidos internos, periostio, paredes arteriales, superficies articulares, hoz del cerebro y tienda del cerebelo. Los estímulos que los excitan pueden ser:
El incremento de sensibilidad de los receptores se conoce como hiperalgesia. La adaptación es muy poca o nula, su importancia es que la falta de adaptación hace consciente a la persona de que el estímulo causante de la lesión tisular sigue actuando.
VELOCIDAD DE LESION TISULAR ESTIMULA EL DOLOR.
La persona comienza a sentir dolor cuando la temperatura alcanza los 45˚C.
La lesión tisular libera químicos que estimulan el dolor.
La isquemia tisular (interrupción del flujo sanguíneo) causa dolor, por la acumulación de grandes acumulaciones de ácido láctico.
El espasmo muscular también, porque aumenta el metabolismo del tejido, lo que incrementa la isquemia relativa, creando condiciones ideales para la liberación de sustancias químicas inductoras de dolor.
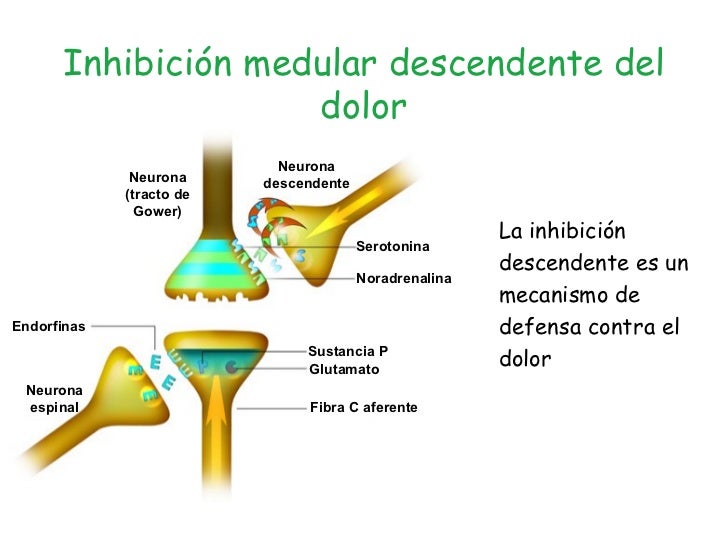
SISTEMA DE SUPRESION DEL DOLOR. Se debe a la capacidad del propio encéfalo para suprimir la entrada de impulsos dolorosos al Sistema Nervioso mediante la activación de un sistema de control llamado Sistema de Analgesia, formado por:
1) Sustancia gris perisilviana y las áreas periventriculares del mesencéfalo 2) Núcleo magno del rafe y núcleo reticular paragigantocelular 3) Complejo inhibidor del dolor situado en las astas posteriores de la célula.
El sistema bloquea las señales de dolor a su entrada en la médula espinal. En el sistema de analgesia intervienen distintas sustancias transmisoras:
Sustancias Opiáceas Importantes:
β-Endorfina.
Met-Encefalina.
Leu-Encefalina.
Dinorfina.
Todas derivan de la proopiomelanocortina, proencefalina y prodinorfina. Existen dos tratamientos del dolor, la inhibición por impulsos sensitivos táctiles (frotar la piel y acupuntura), y el tratamiento por estimulación eléctrica (electrodos).
DOLOR REFERIDO. El dolor se percibe en una parte del cuerpo muy alejada de los tejidos donde se origina. El dolor comienza en una víscera y es referido a una región de la superficie corporal.
DOLOR VISCERAL Dolor procedente de las distintas vísceras abdominales o torácicas. Dolor superficial y dolor visceral. Las causas del dolor visceral verdadero puede ser isquemia de los tejidos viscerales. Todos los dolores viscerales verdaderos que se originan en cavidad torácica o abdominal, se transmiten a través de fibras nerviosas que conducen dolor y discurren con los nervios simpáticos. Fibras tipo C. 1) Isquemia. 2) Espasmo de una víscera hueca (calambres). 3) Sobredistensión de una víscera hueca.
VISCERAS INSENSIBLES (Parénquima Hepático y Alvéolos pulmonares).
Pero la cápsula del hígado es sumamente sensible a traumatismos directos como la distensión, y también los conductos biliares.
Los alvéolos pulmonares son insensibles, mientras que los bronquios y pleura parietal muestran gran sensibilidad.
DOLOR PARIETAL POR LA LESION DE UNA VISCERA. Cuando una enfermedad afecta una víscera, el proceso patológico se extiende a la hoja parietal del peritoneo, pleura o pericardio.
LOCALIZACION DEL DOLOR VISCERAL. Se origina de distintas vísceras, difícil de localizar. Se pueden transmitir por la vía visceral verdadera (referidas a zonas superficiales, muchas veces alejadas del órgano que duele) o por la vía parietal (localizadas directamente por la zona dolorosa, dolor torácico y abdominal).
ALTERACIONES DEL DOLOR.
Hiperalgesia
Síndrome Talámico
Herpes Zoster
Tic Doloroso
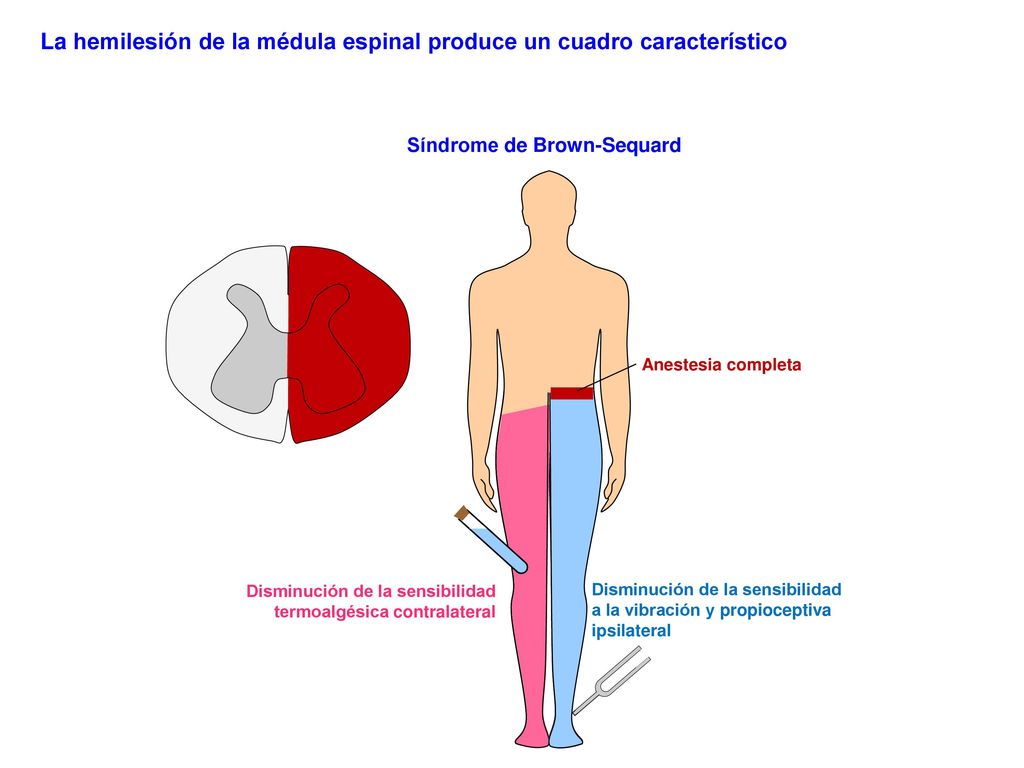
Síndrome de Brown-Séquard
CEFALEAS.
La cefalea constituye un tipo de dolor referido de la superficie de la cabeza que procede de estructuras profundas.
CEFALEA INTRACRANEAL. Se refiere a regiones como la región occipital. 1) Cefalea de Meningitis. 2) Cefalea por Hipotensión de LCR. 3) Jaqueca (Vascular). 4) Cefalea Alcohólica. 5) Cefalea por Estreñimiento.
CEFALEA EXTRACRANEAL. 1) Cefalea por espasmo muscular (Tensión o Estrés). 2) Cefalea por irritación de estructuras nasales y paranasales. 3) Cefalea por trastornos oculares.
SENSACIONES TERMICAS.
Los seres humanos perciben distintos grados de frío y calor:
Frío de congelación.
Frío.
Fresco.
Temperatura normal.
Tibieza.
Calor.
Calor Abrasador.
RECEPTORES. Fibras de dolor estimuladas por frío, fibras de frío, fibra de calor, fibra de dolor estimulada por calor. Es importante mencionar la respuesta a cambios de temperatura, y la adaptación casi total a la temperatura.
. Cuando la temperatura sube de +10 a 15 °C, cesan los impulsos dolorosos de frío, pero empiezan a verse estimulados los receptores para el frío, alcanzando su máximo en torno a los 24 °C y apagándose un poco por encima de 40 °C. Por encima de unos 30 °C, comienzan a estimularse los receptores para el calor, pero también se extinguen más o menos sobre los 49 °C. Finalmente, alrededor de los 45 °C, las fibras para el dolor por el calor empiezan a ser estimuladas por esta situación y, paradójicamente, de nuevo también algunas de las fibras para el frío, tal vez debido a la lesión de sus terminaciones correspondientes ocasionada por el excesivo calor.
TRANSMISION DE LAS SEÑALES AL SISTEMA NERVIOSO.
En general, las señales térmicas se transmiten por vías paralelas a las que siguen las señales dolorosas. Al entrar en la médula espinal, ascienden o descienden unos cuantos segmentos por el fascículo de Lissauer y después terminan sobre todo en las láminas I, II y III de las astas dorsales: las mismas que en el caso del dolor. Después de un cierto grado de procesamiento en una neurona medular o en más, las señales se incorporan a fibras térmicas ascendentes largas que cruzan hacia el fascículo sensitivo anterolateral opuesto y acaban en: 1) la formación reticular del tronco del encéfalo, y 2) el complejo ventrobasal del tálamo.
usculo liso no contiene estrías, es involuntario y está formado por fibras mucho menores, de 1 a 5 mm de diámetro y de sólo 20 a 500 mm de longitud, y aunque tiene las misma fuerzas de atracción entre los filamentos de miosina y actina que el musculo esquelético, tiene algo más que lo caracteriza.
Tipos de músculo liso
Diferencias de los diferentes tipos de musculo liso:
dimensiones físicas
organización en fascículos o láminas
respuesta a diferentes tipos de estímulos
características de la inervación
función.
Sin embargo, en aras de la simplicidad, el músculo liso en general se puede dividir en dos tipos principales
músculo liso multiunitario
músculo liso unitario (o monounitario).
Músculo liso multiunitario
Está formado por fibras musculares lisas separadas y discretas, cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa, la superficie externa de estas fibras, está cubierta por una capa delgada de sustancia similar a una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí.
Ejemplos
El músculo ciliar del ojo
el músculo del iris del ojo
los músculos piloerectores que producen la erección del pelo cuando los estimula el sistema nervioso simpático
Músculo liso unitario (músculo liso sincitial o músculo liso visceral)
Una masa de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad.
Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra, de modo que los potenciales de acción o el flujo iónico simple sin potenciales de acción puede viajar desde una fibra a otra y hacer que las fibras musculares se contraigan simultáneamente.
Se encuentra en la pared de la mayor parte de las vísceras del cuerpo.
Ejemplo
el aparato digestivo
las vías biliares
los uréteres,
el útero
vasos sanguíneos
Mecanismo contráctil en el músculo liso
Base química de la contracción del músculo liso
El músculo liso contiene filamentos tanto de actina como de miosina, que tienen características químicas similares a los filamentos de actina y miosina del músculo esquelético.
Los filamentos de actina y miosina del músculo liso interactúan entre sí de manera muy similar a como lo hacen en el músculo esquelético. Además, el proceso contráctil es activado por los iones calcio, y el trifosfato de adenosina (ATP) se degrada a difosfato de adenosina (ADP) para proporcionar la energía para la contracción. Pero también tiene diferencias importantes entre la organización física del músculo liso y la del músculo esquelético, así como diferencias en el acoplamiento excitación-contracción, el control del proceso contráctil por los iones calcio, la duración de la contracción y la cantidad de energía necesaria.
Base física de la contracción del músculo liso
Debido a que el musculo liso no contiene estrías, contiene cuerpos densos algunos de estos cuerpos están unidos a la membrana celular, otros están dispersos en el interior de la célula y algunos de los cuerpos densos de la membrana de células adyacentes están unidos entre sí por puentes proteicos intercelulares.
La fuerza de contracción se transmite de unas células a otras principalmente a través de estos enlaces. Hay otra diferencia: la mayor parte de los filamentos de miosina tiene lo que se denomina puentes cruzados «lateropolares», dispuestos de tal manera que los puentes de un lado basculan en una dirección y los del otro lado basculan en la dirección opuesta. Esto permite que la miosina tire de un filamento de actina en una dirección en un lado a la vez que simultáneamente tira de otro filamento de actina en la dirección opuesta en el otro lado. La utilidad de esta organización es que permite que las células musculares lisas se contraigan hasta el 80% de su longitud, en lugar de estar limitadas a menos del 30%, como ocurre en el músculo esquelético.
Comparación de la contracción del músculo liso con la contracción del músculo estriado
Ciclado lento de los puentes cruzados de miosina
La rapidez del ciclado de los puentes transversos de miosina en el músculo liso (es decir, su unión a la actina, su posterior liberación de la actina y su nueva unión para el siguiente ciclo) es mucho más lenta que en el músculo esquelético, la fracción de tiempo que los puentes cruzados permanecen unidos a los filamentos de actina, que es un factor importante que determina la fuerza de la contracción, está muy aumentada en el músculo liso.
Baja necesidad de energía para mantener la contracción del músculo liso
La baja utilización de energía por el músculo liso es muy importante para la economía energética global del cuerpo, porque órganos como los intestinos, la vejiga urinaria, la vesícula biliar y otras vísceras con frecuencia mantienen una contracción muscular tónica casi indefinidamente.
Lentitud del inicio de la contracción y relajación del tejido muscular liso total
Un tejido muscular liso típico comienza a contraerse de 50 a 100 ms después de ser excitado, alcanza la contracción completa aproximadamente 0,5 s después, y después la fuerza contráctil disminuye en 1 a 2 segundos más, dando un tiempo total de contracción de 1 a 3 s. Pero como hay tantos tipos de músculo liso, la contracción de algunos tipos puede ser tan corta como 0,2 s o tan larga como 30 s.
El inicio lento de la contracción del músculo liso, así como su contracción prolongada, está producido por la lentitud de la unión y la separación de los puentes cruzados a los filamentos de actina.
La fuerza máxima de contracción muscular es a menudo mayor en el músculo liso que en el músculo esquelético.
La fuerza máxima de contracción del músculo liso es con frecuencia mayor que la del músculo esquelético, hasta 4 a 6 kg/cm2 de área transversal para el músculo liso. Esta gran fuerza de la contracción del músculo liso se debe al período prolongado de unión de los puentes cruzados de miosina a los filamentos de actina.
El mecanismo de «cerrojo» facilita el mantenimiento prolongado de las contracciones del músculo liso
La energía que se consume para mantener la contracción con frecuencia es minúscula, a veces tan sólo 1/300 de la energía necesaria para una contracción sostenida y comparable del músculo esquelético. Esto se denomina mecanismo de «cerrojo». La importancia del mecanismo de cerrojo es que permite mantener una contracción tónica prolongada en el músculo liso durante horas con un bajo consumo de energía. Es necesaria una señal excitadora continua baja procedente de las fibras nerviosas o de fuentes hormonales.
Tensión-relajación del músculo liso
Una característica importante del músculo liso es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado.
Cuando se produce una reducción súbita de volumen, la presión disminuye drásticamente al principio, aunque después aumenta en un plazo de otros pocos segundos o minutos hasta el nivel original o casi hasta el mismo. Estos fenómenos se denominan tensión-relajación y tensión-relajación inversa, permiten que un órgano hueco mantenga aproximadamente la misma presión en el interior de su luz a pesar de grandes cambios de volumen a largo plazo.
Regulación de la contracción por los iones calcio
El estímulo que inicia la mayor parte de las contracciones del músculo liso es un aumento de los iones calcio en el medio intracelular. Este aumento puede estar producido en diferentes tipos de músculo liso por la estimulación nerviosa de las fibras de músculo liso, por estimulación hormonal, por distensión de la fibra o incluso por cambios del ambiente químico de la fibra
El músculo liso no contiene troponina, la proteína reguladora que es activada por los iones calcio para producir la contracción del músculo esquelético pero contienen una gran cantidad de otra proteína reguladora denominada calmodulina, activa los puentes cruzados de miosina.
Esta activación y la posterior contracción se producen según la siguiente secuencia:
Los iones calcio se unen a la calmodulina
El complejo calmodulina-calcio se une después a la miosina cinasa de cadena ligera, que es una enzima fosforiladora, y la activa.
Una de las cadenas ligeras de cada una de las cabezas de miosina, denominada cabeza reguladora, se fosforila en respuesta a esta miosina cinasa. Cuando esta cadena no está fosforilada no se produce el ciclo de unión-separación de la cabeza de miosina con el filamento de actina, pero cuando la cadena reguladora está fosforilada la cabeza tiene la capacidad de unirse repetitivamente al filamento de actina y de avanzar a través de todo el proceso de ciclado de «tirones» intermitentes, al igual que ocurre en el músculo esquelético, produciendo de esta manera la contracción muscular.
La miosina fosfatasa
El tiempo necesario para la relajación de la contracción muscular está determinado en gran medida por la cantidad de miosina fosfatasa activa en la célula.
Posible mecanismo de regulación del fenómeno de cerrojo
Este fenómeno permite el mantenimiento a largo plazo del tono en muchos órganos que tienen músculo liso sin un gran gasto de energía.
Cuando las enzimas miosina cinasa y miosina fosfatasa están intensamente activadas las dos, la frecuencia de ciclado de las cabezas de miosina y la velocidad de contracción son elevadas. Después, cuando disminuye la activación de las enzimas, lo hace también la frecuencia de ciclado, pero al mismo tiempo la desactivación de estas enzimas permite que las cabezas de miosina permanezcan unidas al filamento de actina durante una proporción cada vez mayor del período de ciclado, como el número de cabezas unidas a la actina determina la fuerza estática de la contracción, se mantiene, o cierra, la tensión.
Control nervioso y hormonal de la contracción del músculo liso
La contracción del músculo liso puede ser estimulada por múltiples tipos de señales: señales nerviosas, estimulación hormonal, distensión del músculo y otros diversos estímulos. El principal motivo de esta diferencia es que la membrana del músculo liso contiene muchos tipos de proteínas receptoras que pueden iniciar el proceso contráctil.
Uniones neuromusculares del músculo liso
Anatomía fisiológica de las uniones neuromusculares del músculo liso
Las uniones neuromusculares del tipo muy estructurado que se encuentran en las fibras del músculo esquelético no aparecen en el músculo liso. Por el contrario, las fibras nerviosas autónomas que inervan el músculo liso generalmente se ramifican de manera difusa encima de una lámina de fibras musculares, estas fibras no hacen contacto directo con la membrana de las células de las fibras musculares lisas, sino que forman las denominadas uniones difusas que secretan su sustancia transmisora hacia el recubrimiento de matriz del músculo liso, con frecuencia a una distancia de varios nanómetros a varios micrómetros de las células musculares; después la sustancia transmisora difunde hacia las células. La excitación muscular viaja desde esta capa externa hacia las capas internas por conducción de los potenciales de acción en la masa muscular o mediante difusión adicional de la sustancia transmisora.
La mayor parte de los axones terminales delgados tiene múltiples varicosidades distribuidas a lo largo de sus ejes. En estos puntos se interrumpen las células de Schwann que rodean a los axones, de modo que se puede secretar la sustancia transmisora a través de las paredes de las varicosidades. Al contrario de las vesículas de las uniones del músculo esquelético, que siempre contienen acetilcolina, las vesículas de las terminaciones de las fibras nerviosas autónomas contienen acetilcolina en algunas fibras y noradrenalina en otras.
Sustancias transmisoras excitadoras e inhibidoras secretadas en la unión neuromuscular del músculo liso.
acetilcolina y noradrenalina
La acetilcolina es una sustancia transmisora excitadora de las fibras musculares lisas en algunos órganos y un transmisor inhibidor en el músculo liso de otros órganos. Cuando la acetilcolina excita una fibra, la noradrenalina habitualmente la inhibe. Por el contrario, cuando la acetilcolina inhibe una fibra, la noradre nalina habitualmente la excita.
Tanto la acetilcolina como la noradrenalina excitan o inhiben el músculo liso uniéndose en primer lugar a una proteína receptora de la superficie de la membrana de la célula muscular. Algunas de las proteínas receptoras son receptores excitadores, mientras que otras son receptores inhibidores.
Potenciales de membrana y potenciales de acción en el músculo liso
Potencial de membrana
El voltaje cuantitativo del potencial de membrana del músculo liso depende de la situación momentánea del músculo.
Estado de reposo normal el potencial intracelular es habitualmente de aproximadamente –50 a –60 mV.
Potencial de acción
Los potenciales de acción se producen en el músculo liso
unitario (como el músculo visceral) de la misma forma que en el músculo esquelético.
Los potenciales de acción del músculo liso visceral se producen en una de dos formas:
potenciales en espiga
potenciales de acción con meseta
Potenciales en espiga
La duración de este tipo de potencial de acción es de 10 a 50 ms, se pueden generar de muchas maneras, por ejemplo mediante estimulación eléctrica, por la acción de hormonas sobre el músculo liso, por la acción de sustancias transmisoras procedentes de las fibras nerviosas, entre otros.
Potenciales de acción con meseta
El inicio de este potencial de acción es similar al del potencial en espiga típico, la repolarización se retrasa durante varios cientos hasta 1.000 ms (1 segundo), esta meseta puede ser responsable de la contracción prolongada que se produce en algunos tipos de músculo liso, como el uréter, el útero en algunas situaciones y ciertos tipos de músculo liso vascular.
Los canales de calcio
El sodio participa poco en la generación del potencial de acción en la mayor parte del músculo liso, el flujo de iones calcio hacia el interior de la fibra es el principal responsable del potencial de acción
Los canales de calcio se abren muchas veces más lentos que los canales de sodio, y también permanecen abiertos mucho más tiempo. Esto explica en gran medida los prolongados potenciales de acción en meseta de algunas fibras musculares lisas.
Los potenciales de onda lenta en el músculo liso unitario
Pueden conducir a la generación espontánea de potenciales de acción, los potenciales de acción se originan en las propias células musculares lisas sin ningún estímulo extrínseco. La propia onda lenta no es el potencial de acción. Es decir, no es un proceso autorregenerativo que se propaga progresivamente a lo largo de las membranas de las fibras musculares, sino que es una propiedad local de las fibras musculares lisas que forman la masa muscular.
Cuando son lo suficientemente intensas, pueden iniciar potenciales de acción. Las ondas lentas en sí mismas no pueden producir la contracción muscular. No obstante, cuando el máximo del potencial de la onda lenta negativa en el interior de la membrana celular aumenta en dirección positiva desde –60 hasta aproximadamente –35 mV se produce un potencial de acción que se propaga a lo largo de la masa muscular y se produce la contracción.
Por tanto, las ondas lentas se denominan ondas marcapasos.
Excitación del músculo liso visceral por distensión muscular
Cuando el músculo liso visceral (unitario) es distendido lo suficiente, se generan potenciales de acción espontáneos:
Los potenciales de onda lenta normales
la disminución de la negatividad global del potencial de membrana que produce la propia distensión.
Despolarización del músculo liso multiunitario sin potenciales de acción
Las terminaciones nerviosas secretan acetilcolina en el caso de algunos músculos lisos multiunitarios y noradrenalina en el caso de otros, en ambos casos, las sustancias transmisoras producen despolarización de la membrana del músculo liso, y esto a su vez produce la contracción.
No se producen potenciales de acción; el motivo es que las fibras son demasiado pequeñas para generar un potencial de acción, aunque las células musculares lisas pequeñas, incluso sin potencial de acción, la despolarización local (denominada potencial de la unión) que produce la propia sustancia transmisora nerviosa se propaga electrotónicamente en toda la fibra y es lo único necesario para producir la contracción muscular.
Los efectos de los factores tisulares locales y las hormonas
Determinan la contracción del músculo liso sin potenciales de acción, dos tipos de factores estimulantes no nerviosos y no relacionados con el potencial de acción que participan con frecuencia son:
factores químicos tisulares locales
varias hormonas.
Contracción del músculo liso en respuesta a factores químicos tisulares locales
El músculo liso es muy contráctil y responde rápidamente a los cambios de las condiciones químicas locales del líquido intersticial circundante, un potente sistema de control de retroalimentación local controla el flujo sanguíneo a la zona tisular local. Algunos de los factores de control específicos son los siguientes:
La ausencia de oxígeno en los tejidos locales produce relajación del músculo liso y, por tanto, vasodilatación.
El exceso de anhídrido carbónico produce vasodilatación.
El aumento de la concentración de iones hidrógeno produce vasodilatación.
Efectos de las hormonas sobre la contracción del músculo liso
Una hormona produce contracción del músculo liso cuando la membrana de la célula muscular contiene receptores excitadores activados por hormonas para esa hormona. Por el contrario, la hormona produce inhibición si la membrana contiene receptores inhibidores para ella en lugar de receptores excitadores.
Mecanismos de la excitación o la inhibición del músculo liso por hormonas o por factores tisulares locales
La contracción o la inhibición del músculo liso es iniciada por hormonas que no producen directamente ningún cambio en el potencial de membrana, para inhibir la contracción se activar la enzima adenilato ciclasa o guanilato ciclasa de la membrana celular; las porciones de los receptores que sobresalen hacia el interior de las células están acopladas con estas enzimas, dando lugar a la formación de monofosfato cíclico de adenosina (AMPc) o monofosfato cíclico de guanosina (GMPc), denominados segundos mensajeros.
El AMPc y el GMPc tienen muchos efectos, uno de los cuales es modificar el grado de fosforilación de varias enzimas que inhiben indirectamente la contracción. Se activa la bomba que mueve iones calcio desde el sarcoplasma hacia el retículo sarcoplásmico, así como la bomba de la membrana celular que saca iones calcio de la propia célula; estos efectos reducen la concentración de los iones calcio en el sarcoplasma, inhibiendo de esta manera la contracción.
Origen de los iones calcio
Causan la contracción a través de la membrana celular y a partir del retículo sarcoplásmico, el retículo sarcoplásmico, está poco desarrollado en la mayor parte del músculo liso y la mayoría de los iones calcio que producen la contracción entran en la célula muscular desde el líquido extracelular en el momento del potencial de acción o de otro estímulo y la concentración de iones calcio en el líquido extracelular es superior a 10–3 molar, en comparación con menos de 10–7 molar en el interior de la célula muscular lisa; esto produce una difusión rápida de los iones calcio hacia el interior de la célula desde el líquido extracelular cuando se abren los canales de calcio. El tiempo necesario para que se produzca esta difusión es en promedio de entre 200 y 300 ms y se denomina período de latencia antes de que comience la contracción.
Función del retículo sarcoplásmico del músculo liso
Pequeñas invaginaciones de la membrana celular, denominadas cavéolas, están junto a las superficies de los túbulos sarcoplásmicos, la transmisión de un potencial de acción hacia las cavéolas excita la liberación de iones calcio desde los túbulos sarcoplásmicos próximos de la misma manera que los potenciales de acción de los túbulos transversos del músculo esquelético producen la liberación de iones calcio desde los túbulos sarcoplásmicos longitudinales del músculo esquelético.
Contracción del músculo liso
Depende de la concentración extracelular de los iones calcio, Cuando la concentración de iones calcio en el líquido extracelular disminuye a aproximadamente 1/3 a 1/10 de su valor normal, habitualmente se interrumpe la contracción del músculo liso. Por tanto, la fuerza de la contracción del músculo liso depende mucho de la concentración de iones calcio en el líquido extracelular.
Es necesaria una bomba de calcio para producir la relajación del músculo liso, y esta bombea iones calcio hacia el exterior de la fibra muscular lisa de nuevo hacia el líquido extracelular o hacia el retículo sarcoplásmico, actúa lentamente, una única contracción del músculo liso con frecuencia dura varios segundos en lugar de centésimas a décimas de segundo, como ocurre en el músculo esquelético.
en la mayoría de los músculos esqueléticos, las fibras se extienden a lo largo de toda la longitud del músculo.
El sarcolema es una fina membrana que envuelve a una fibra musculoesquelética. El sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno.Las fibras tendinosas a su vez se agrupan en haces para formar los tendones musculares, que después insertan los músculos en los huesos.
Las miofibrillas están formadas por filamentos de actina y miosina,Cada fibra muscular contiene varios cientos a varios miles de miofibrillas, que se representan en la vista en sección transversal. está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular real
las miofibrillas tengan bandas claras y oscuras alternas. Las bandas claras contienen solo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada
Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada
El disco Z, que está formado por proteínas filamentosas distintas de los filamentos de actina y miosina, atraviesa las miofibrillas y también pasa desde unas miofibrillas a otras, uniéndolas entre sí a lo largo de toda la longitud de la fibra muscular
La porción de la miofibrilla (o de la fibra muscular entera) que está entre dos discos Z sucesivos se denomina sarcomero
Mecanismo general de la contracción muscular
Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.
En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana.
La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular
El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular
Mecanismo molecular de la contracción muscular
La contracción muscular se produce por un mecanismo de deslizamiento de los filamento
Energética de la contracción muscular
La primera fuente de energía que se utiliza para reconstituir el ATP es la sustancia fosfocreatina, que contiene un enlace fosfato de alta energía similar a los enlaces del ATP.
La segunda fuente importante de energía, que se utiliza para reconstituir tanto el ATP como la fosfocreatina, es la «glucólisis» del glucógeno que se ha almacenado previamente en las células musculares
La tercera y última fuente de energía es el metabolismo oxidativo, lo que supone combinar oxígeno con los productos finales de la glucólisis y con otros diversos nutrientes celulares para liberar ATP.
Características de la contracción de todo el músculo
Las contracciones isométricas no acortan el músculo, mientras que las contracciones isotónicas lo acortan a una tensión constante
la contracción muscular es isométrica cuando el músculo no se acorta durante la contracción e isotónica cuando se acorta, pero la tensión del músculo permanece constante durante toda la contracción.
Mecánica de la contracción del músculo esquelético
Unidad motora: todas las fibras musculares inervadas por una única fibra nerviosa
En general, los músculos pequeños que reaccionan rápidamente y cuyo control debe ser exacto tienen más fibras nerviosas para menos fibras musculares (p. ej., tan solo dos o tres fibras musculares por cada unidad motora en algunos de los músculos laríngeos). Por el contrario, los músculos grandes que no precisan un control fino, como el músculo sóleo, pueden tener varios centenares de fibras musculares en una unidad motora
Remodelación del músculo para adaptarse a la función
Todos los músculos del cuerpo se modelan continuamente para adaptarse a las funciones que deben realizar. Se altera su diámetro, su longitud, su fuerza y su vascularización, e incluso se alteran, al menos ligeramente, los tipos de fibras musculares. Este proceso de remodelación con frecuencia es bastante rápido, y se produce en un plazo de pocas semanas
Hipertrofia y atrofia muscular
El aumento de la masa total de un músculo se denomina hipertrofia muscular. Cuando la masa total disminuye, el proceso recibe el nombre de atrofia muscular. Prácticamente toda la hipertrofia muscular se debe a un aumento del número de filamentos de actina y miosina en cada fibra muscular, dando lugar a aumento de tamaño de las fibras musculares individuales; esta situación se denomina hipertrofia de las fibras
Cuando un músculo no se utiliza durante muchas semanas, la velocidad de degradación de las proteínas contráctiles es mucho más rápida que la velocidad de sustitución. Por tanto, se produce atrofia muscular
Hiperplasia de las fibras musculares
En situaciones poco frecuentes de generación extrema de fuerza muscular se ha observado que hay un aumento real del número de fibras musculares (aunque solo en un pequeño porcentaje), además del proceso de hipertrofia de las fibras
La denervación muscular provoca una rápida atrofia
Cuando un músculo pierde su inervación, ya no recibe las señales contráctiles que son necesarias para mantener el tamaño muscular normal. Por tanto, la atrofia comienza casi inmediatamente. Después de aproximadamente 2 meses también comienzan a aparecer cambios degenerativos en las fibras musculares.
El líquido extracelular contiene una gran cantidad de sodio, pero solo una pequeña cantidad de potasio. En el líquido intracelular ocurre lo contrario. El líquido extracelular contiene una gran cantidad de iones cloruro, mientras que el líquido intracelular contiene muy pocos de estos iones. La concentración de fosfatos y de proteínas del líquido intracelular es considerablemente mayor que la del líquido extracelular. Estas diferencias son muy importantes para la vida de la célula.
La membrana celular consiste en una BICAPA lipídica con proteínas de transporte de la membrana celular. Esta membrana está formada casi totalmente por una bicapa lipídica, aunque también contiene grandes números de moléculas proteicas insertadas en los lípidos, muchas de las cuales penetran en todo el grosor de la membrana.
La bicapa lipídica no es miscible con el líquido extracelular ni con el líquido intracelular. Por tanto, constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular. Sin embargo, las sustancias liposolubles pueden penetrar en esta bicapa lipídica y difunden directamente a través de la sustancia lipídica. Las moléculas proteicas de la membrana tienen unas propiedades totalmente diferentes para transportar sustancias. Sus estructuras moleculares interrumpen la continuidad de la bicapa lipídica y constituyen una ruta alternativa a través de la membrana celular. Algunas tienen espacios acuosos en todo el trayecto del interior de la molécula y permiten el movimiento libre de agua, así como de iones o moléculas seleccionados; estas proteínas se denominan proteínas de los canales. Otras, denominadas proteínas transportadoras, se unen a las moléculas o iones que se van a transportar y cambios conformacionales de las moléculas de la proteína desplazan después las sustancias a través de los intersticios de la proteína hasta el otro lado de la membrana. Las proteínas de los canales y las proteínas transportadoras habitualmente son selectivas para los tipos de moléculas o de iones que pueden atravesar la membrana.
«Difusión» frente a «transporte activo»
El transporte a través de la membrana celular, ya sea directamente a través de la bicapa lipídica o a través de las proteínas, se produce mediante uno de dos procesos básicos: difusión o transporte activo.
La difusión se refiere a un movimiento molecular aleatorio de las sustancias molécula a molécula, a través de espacios intermoleculares de la membrana o en combinación con una proteína transportadora. La energía que hace que se produzca la difusión es la energía del movimiento cinético normal de la materia.
El transporte activo se refiere al movimiento de iones o de otras sustancias a través de la membrana en combinación con una proteína transportadora de tal manera que la proteína transportadora hace que la sustancia se mueva contra un gradiente de energía, como desde un estado de baja concentración a un estado de alta concentración. Este movimiento precisa una fuente de energía adicional, además de la energía cinética.
Difusión
Todas las moléculas e iones de los líquidos corporales, incluyendo las moléculas de agua y las sustancias disueltas, están en movimiento constante, de modo que cada partícula se mueve de manera completamente independiente. El movimiento de estas partículas es lo que los físicos llaman «calor» (cuanto mayor sea el movimiento, mayor es la temperatura), y el movimiento nunca se interrumpe salvo a la temperatura de cero absoluto. Cuando una molécula en movimiento, A, se acerca a una molécula estacionaria, B, las fuerzas electrostáticas y otras fuerzas nucleares de la molécula A rechazan a la molécula B, transfiriendo parte de la energía del movimiento de la molécula A a la B. En consecuencia, la molécula B adquiere energía cinética del movimiento, mientras que la molécula A se enlentece, perdiendo parte de su energía cinética.
Difusión a través de la membrana celular
Se divide en dos subtipos:
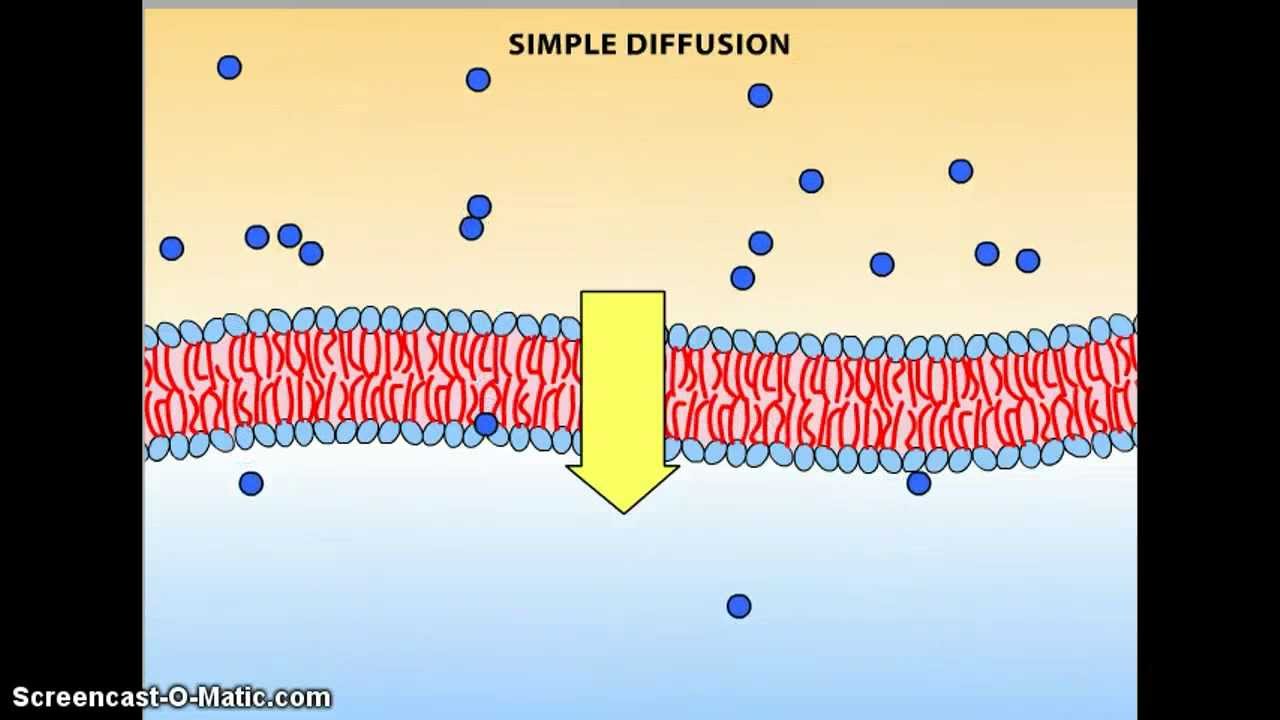
*Difusión simple: el movimiento cinético de las moléculas o de los iones se produce a través de una abertura de la membrana o a través de espacios intermoleculares sin ninguna interacción con las proteínas transportadoras de la membrana. La velocidad de difusión viene determinada por la cantidad de sustancia disponible, la velocidad del movimiento cinético y el número y el tamaño de las aberturas de la membrana a través de las cuales se pueden mover las moléculas o los iones. Se puede producir difusión simple a través de la membrana celular por dos rutas:
1) a través de los intersticios de la bicapa lipídica si la sustancia que difunde es liposoluble
2) a través de canales acuosos que penetran en todo el grosor de la bicapa a través de las grandes proteínas transportadoras.
*Difusión facilitada: precisa la interacción de una proteína transportadora. La proteína transportadora ayuda al paso de las moléculas o de los iones a través de la membrana mediante su unión química con estos y su desplazamiento a través de la membrana de esta manera.
Difusión de sustancias liposolubles a través de la bicapa lipídica
Un factor importante que determina la rapidez con la que una sustancia difunde a través de la bicapa lipídica es la liposolubilidad. La velocidad de difusión de cada una de estas sustancias a través de la membrana es directamente proporcional a su liposolubilidad.
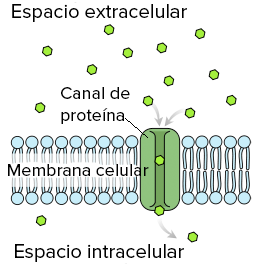
Difusión de agua y de otras moléculas insolubles en lípidos a través de canales proteicos
Aunque el agua es muy insoluble en los lípidos de la membrana, pasa rápidamente a través de los canales de las moléculas proteicas que penetran en todo el espesor de la membrana. Muchas de las membranas celulares del cuerpo contienen «poros» proteicos denominados acuaporinas que permiten selectivamente el rápido paso de agua a través de la membrana celular. Las acuaporinas están muy especializadas, y existen al menos 13 tipos diferentes en las diversas células de los mamíferos. Otras moléculas insolubles en lípidos pueden atravesar los canales de los poros proteicos de la misma manera que las moléculas de agua si son hidrosolubles y de un tamaño lo suficientemente pequeño. Sin embargo, a medida que se hacen mayores su penetración disminuye rápidamente.
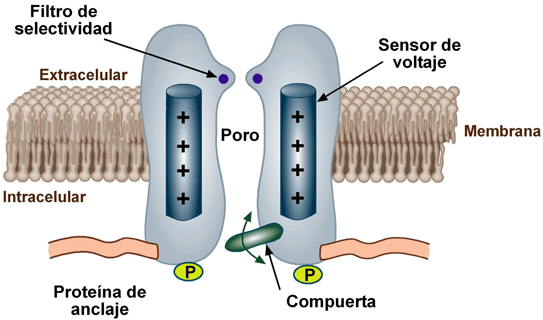
Difusión a través de poros y canales proteicos: permeabilidad selectiva y «activación» de canales
las sustancias se pueden mover mediante difusión simple directamente a lo largo de estos poros y canales desde un lado de la membrana hasta el otro. Los poros están compuestos por proteínas de membranas celulares integrales que forman tubos abiertos a través de la membrana y que están siempre abiertos. Sin embargo, el diámetro de un poro y sus cargas eléctricas proporcionan una selectividad que permite el paso de solo ciertas moléculas a su través. Las acuaporinas tienen un poro estrecho que permite que las moléculas de agua se difundan a través de la membrana en una única fila. El poro es demasiado pequeño para permitir el paso de iones hidratados. Los canales proteicos se distinguen por dos características importantes:
1)con frecuencia son permeables de manera selectiva a ciertas sustancias.
2) muchos de los canales se pueden abrir o cerrar por compuertas que son reguladas por señales eléctricas (canales activados por el voltaje) o sustancias químicas que se unen a las proteínas de canales (canales activados por ligandos).
Permeabilidad selectiva de los canales proteicos
Muchos de los canales proteicos son muy selectivos para el transporte de uno o más iones o moléculas específicos. Esta selectividad se debe a las características del propio canal, como su diámetro, su forma y la naturaleza de las cargas eléctricas y enlaces químicos que están situados a lo largo de sus superficies internas.
Se cree que existen diferentes filtros de selectividad que determinan, en gran medida, la especificidad de los diversos canales para cationes o aniones o para iones determinados, como sodio (Na+), potasio (K+) y calcio (Ca++), que consiguen acceder a los canales. Un ejemplo de los canales proteicos más importantes, el denominado canal del sodio, mide solo de 0,3 a 0,5 nm de diámetro, aunque, lo que es más importante, las superficies internas de este canal están revestidas con aminoácidos que tienen una carga intensamente negativa. Estas cargas negativas intensas pueden arrastrar pequeños iones sodio deshidratados hacia el interior de estos canales, realmente separando los iones sodio de las moléculas de agua que los hidratan. Una vez que están en el canal, los iones sodio difunden en una u otra dirección según las leyes habituales de la difusión. Así, el canal del sodio es altamente selectivo para el paso de iones sodio.
Activación de los canales proteicos
La activación de los canales proteicos proporciona un medio para controlar la permeabilidad iónica de los canales. Se piensa que algunas de las compuertas son realmente extensiones similares a una compuerta de la molécula de la proteína transportadora, que pueden cerrar la abertura del canal o se pueden alejar de la apertura por un cambio conformacional de la forma de la propia molécula proteica. La apertura y el cierre de las compuertas están controlados de dos maneras principales:
1. Activación por el voltaje: la conformación molecular de la compuerta o de sus enlaces químicos responde al potencial eléctrico que se establece a través de la membrana celular.
2. Activación química (por ligando): Las compuertas de algunos canales proteicos se abren por la unión de una sustancia química (un ligando) a la proteína, que produce un cambio conformacional o un cambio de los enlaces químicos de la molécula de la proteína que abre o cierra la compuerta. Uno de los casos más importantes de activación química es el efecto de la acetilcolina sobre el denominado canal de la acetilcolina. La acetilcolina abre la compuerta de este canal, dando lugar a la apertura de un poro de carga negativa que permite que lo atraviesen moléculas sin carga o iones positivos menores de este diámetro. Esta compuerta es muy importante para la transmisión de las señales nerviosas desde una célula nerviosa a otra y desde las células nerviosas a las células musculares para producir la contracción muscular.
La difusión facilitada necesita proteínas transportadoras de membrana
La difusión facilitada también se denomina difusión mediada por un transportador porque una sustancia que se transporta de esta manera difunde a través de la membrana con la ayuda de una proteína transportadora específica para contribuir al transporte. Es decir, el transportador facilita la difusión de la sustancia hasta el otro lado.
La difusión facilitada difiere de la difusión simple en la siguiente característica importante: aunque la velocidad de la difusión simple a través de un canal abierto aumenta de manera proporcional a la concentración de la sustancia que difunde, en la difusión facilitada la velocidad de difusión se acerca a un máximo, denominado Vmáx, a medida que aumenta la concentración de la sustancia que difunde. Entre las numerosas sustancias que atraviesan las membranas celulares mediante difusión facilitada están la glucosa y la mayoría de los aminoácidos. En el caso de la glucosa se han descubierto en varios tejidos al menos 14 miembros de una familia de proteínas de membrana (denominadas GLUT) que transportan moléculas de glucosa. Algunas de estas GLUT también pueden transportar otros monosacáridos que tienen estructuras similares a la glucosa, entre ellos la galactosa y la fructosa. Una de ellas, el transportador de glucosa 4 (GLUT4), es activada por insulina, lo que puede aumentar la velocidad de la difusión facilitada de la glucosa hasta 10 a 20 veces en tejidos sensibles a la insulina.
Factores que influyen en la velocidad neta de difusión
La velocidad neta de difusión es proporcional a la diferencia de concentración a través de una membrana
La velocidad a la que la sustancia difunde hacia dentro es proporcional a la concentración de las moléculas en el exterior, porque esta concentración determina cuántas moléculas chocan contra el exterior de la membrana cada segundo. Por el contrario, la velocidad a la que las moléculas difunden hacia afuera es proporcional a su concentración en el interior de la membrana. Por tanto, la velocidad de difusión neta hacia el interior de la célula es proporcional a la concentración en el exterior menos la concentración en el interior.
Efecto de una diferencia de presión a través de la membrana
En ocasiones se produce una gran diferencia de presión entre los dos lados de una membrana permeable. Esta diferencia de presión se produce, por ejemplo, en la membrana capilar sanguínea de todos los tejidos del cuerpo. La presión es aproximadamente 20 mmHg mayor en el interior del capilar que en el exterior. La presión realmente significa la suma de todas las fuerzas de las diferentes moléculas que chocan contra una unidad de superficie en un momento dado. Por tanto, al tener una presión mayor en un lado de la membrana que en el otro, la suma de todas las fuerzas de las moléculas que chocan con los canales de ese lado de la membrana es mayor que en el otro lado. En la mayor parte de los casos esta situación se debe a que hay un mayor número de moléculas que choca cada segundo contra la membrana en un lado que contra la del otro lado. La consecuencia es que se dispone de mayores cantidades de energía para producir el movimiento neto de moléculas desde el lado de presión elevada hacia el lado de presión baja.
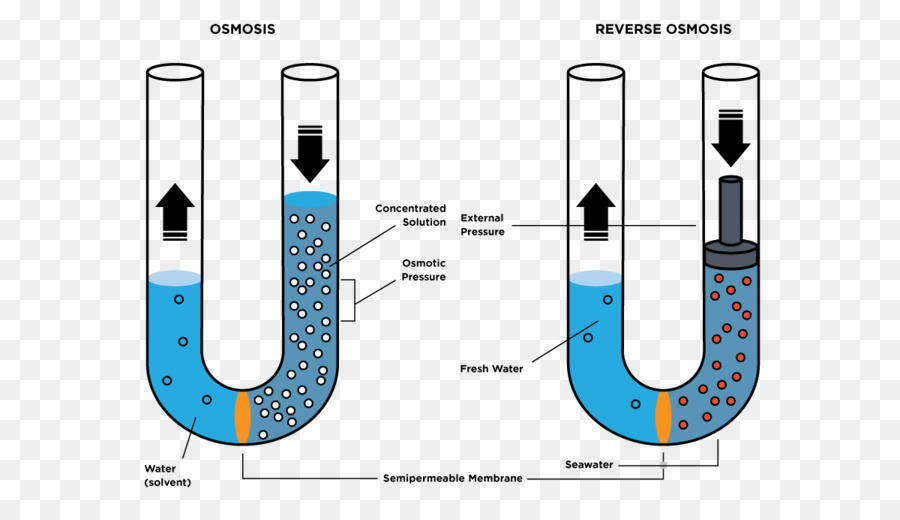
Ósmosis a través de membranas con permeabilidad selectiva: «difusión neta» de agua
La sustancia más abundante que difunde a través de la membrana celular es el agua. en ciertas condiciones se puede producir una diferencia de concentración del agua a través de la membrana. Cuando se produce esto, tiene lugar un movimiento neto de agua a través de la membrana celular, haciendo que la célula se hinche o que se contraiga, dependiendo de la dirección del movimiento del agua. Este proceso de movimiento neto del agua que se debe a la producción de una diferencia de la concentración del agua se denomina ósmosis. Para ilustrar la ósmosis debemos asumir las condiciones en la que hay agua pura a un lado de la membrana celular y una solución de cloruro sódico en el otro lado. Las moléculas de agua atraviesan la membrana celular con facilidad, mientras que los iones sodio y cloruro pasan solo con dificultad. Sin embargo, la presencia del sodio y del cloruro ha desplazado parte de las moléculas de agua del lado de la membrana en el que están presentes estos iones y, por tanto, ha reducido la concentración de moléculas de agua a una concentración menor que la del agua pura. En consecuencia, más moléculas de agua chocan contra los canales del lado izquierdo, en el que hay agua pura, que en el lado derecho, en el que se ha reducido la concentración de agua. Así, se produce un movimiento neto de agua desde la izquierda hacia la derecha, es decir, se produce ósmosis desde el agua pura hacia la solución de cloruro sódico.
Presión osmótica
La cantidad de presión necesaria para detener la ósmosis se denomina presión osmótica de la solución de cloruro sódico. El principio de una diferencia de presión que se opone a la ósmosis, que muestra una membrana con permeabilidad selectiva que separa dos columnas de líquido, una que contiene agua pura y otra que contiene una solución de agua y de cualquier soluto que no penetra en la membrana. La ósmosis de agua desde la cámara B hacia la cámara A hace que los niveles de las columnas de líquido se separen cada vez más, hasta que finalmente se produzca una diferencia de presión entre los dos lados de la membrana que sea lo suficientemente grande como para oponerse al efecto osmótico. Esta diferencia de presión a través de la membrana en este punto es igual a la presión osmótica de la solución que contiene el soluto no difusible.
Importancia del número de partículas osmóticas (concentración molar) en la determinación de la presión osmótica
La presión osmótica que ejercen las partículas de una solución, ya sean moléculas o iones, está determinada por el número de partículas por unidad de volumen del líquido, no por la masa de las partículas. La razón de esto es que todas las partículas de una solución, independientemente de su masa, ejercen, en promedio, la misma cantidad de presión contra la membrana.
«Osmolalidad»: el osmol
Para expresar la concentración de una solución en función del número de partículas se utiliza la unidad denominada osmol en lugar de los gramos. Un osmol es el peso molecular-gramo de un soluto osmóticamente activo. Por tanto, 180 g de glucosa, que es el peso molecular-gramos de la glucosa, son equivalentes a un osmol de glucosa porque la glucosa no se disocia en iones. Si un soluto se disocia en dos iones, un peso molecular-gramo del soluto se convertirá en dos osmoles porque el número de partículas osmóticamente activas es ahora el doble que en el caso del soluto no disociado. Se dice que una solución que tiene 1 osmol de soluto disuelto por cada kilogramo de agua tiene una osmolalidad de 1 osmol por kilogramo, y una solución que tiene 1/1.000 osmoles disueltos por kilogramo tiene una osmolalidad de 1 mosmol por kilogramo. La osmolaridad normal de los líquidos extracelular e intracelular es de aproximadamente 300 mosmol por kilogramo de agua.
osmolaridad
Osmolaridad es la concentración osmolar expresada en osmoles por litro de solución en lugar de osmoles por kilogramo de agua. Aunque en sentido estricto son los osmoles por kilogramo de agua (osmolalidad) los que determinan la presión osmótica, para las soluciones diluidas como las que se encuentran en el cuerpo las diferencias cuantitativas entre la osmolaridad y la osmolalidad son menores del 1%. Como es mucho más práctico medir la osmolaridad que la osmolalidad, esta es la práctica habitual en casi todos los estudios fisiológicos.
«Transporte activo» de sustancias a través de las membranas
Cuando una membrana celular transporta moléculas o iones «contra corriente» contra un gradiente de concentración (o «contra corriente» contra un gradiente eléctrico o de presión), el proceso se denomina transporte activo. Diferentes sustancias que se transportan activamente a través de al menos algunas membranas celulares incluyen los iones sodio, potasio, calcio, hierro, hidrógeno, cloruro, yoduro y urato, diversos azúcares diferentes y la mayoría de los aminoácidos.
Transporte activo primario y transporte activo secundario
El transporte activo se divide en dos tipos según el origen de la energía que se utiliza para facilitar el transporte:
1.-transporte activo primario: la energía procede directamente de la escisión del trifosfato de adenosina (ATP) o de algún otro compuesto de fosfato de alta energía.
2.-transporte activo secundario: la energía procede secundariamente de la energía que se ha almacenado en forma de diferencias de concentración iónica de sustancias moleculares o iónicas secundarias entre los dos lados de una membrana celular, que se generó originalmente mediante transporte activo primario.
En ambos casos el transporte depende de proteínas transportadoras que penetran a través de la membrana celular, al igual que en la difusión facilitada. Sin embargo, en el transporte activo la proteína transportadora funciona de manera diferente al transportador de la difusión facilitada porque es capaz de impartir energía a la sustancia transportada para moverla contra el gradiente electroquímico.
Transporte activo primario
La bomba sodio-potasio transporta iones sodio hacia el exterior de las células e iones potasio hacia el interior
Entre las sustancias que se transportan mediante transporte activo primario están el sodio, el potasio, el calcio, el hidrógeno, el cloruro y algunos otros iones. El mecanismo de transporte activo que se ha estudiado con mayor detalle es la bomba sodio-potasio (Na+-K+), que es el proceso de transporte que bombea iones sodio hacia fuera a través de la membrana celular de todas las células y al mismo tiempo bombea iones potasio desde el exterior hacia el interior. Esta bomba es responsable de mantener las diferencias de concentración de sodio y de potasio a través de la membrana celular, así como de establecer un voltaje eléctrico negativo en el interior de las células.
La proteína transportadora es un complejo formado por dos proteínas globulares distintas: una de mayor tamaño denominada subunidad α, que tiene un peso molecular de aproximadamente 100.000, y una más pequeña denominada subunidad β, que tiene un peso molecular de aproximadamente 55.000. Aunque se desconoce la función de la proteína de menor tamaño (excepto que podría anclar el complejo proteico a la membrana lipídica), la proteína de mayor tamaño tiene tres características específicas que son importantes para el funcionamiento de la bomba:
1. Tiene tres puntos receptores para la unión de iones sodio en la porción de la proteína que protruye hacia el interior de la célula.
2. Tiene dos puntos receptores para iones potasio en el exterior.
3. La porción interior de esta proteína cerca de los puntos de unión al sodio tiene actividad adenosina trifosfatasa (ATPasa).
Cuando dos iones potasio se unen al exterior de la proteína transportadora y tres iones sodio se unen al interior se activa la función ATPasa de la proteína. La activación de la función ATPasa conduce a una escisión de una molécula de ATP, que se divide en difosfato de adenosina (ADP) y libera un enlace de energía de fosfato de alta energía. Se piensa que esta energía liberada produce un cambio químico y conformacional en la molécula transportadora proteica, transportando los tres iones sodio hacia el exterior y los dos iones potasio hacia el interior.
La bomba Na+-K+ es importante para controlar el volumen celular
Una de las funciones más importantes de la bomba Na+-K+ es controlar el volumen de todas las células. Sin la función de esta bomba la mayoría de las células del cuerpo se hincharían hasta explotar. El mecanismo para controlar el volumen es el siguiente: en el interior de la célula hay grandes cantidades de proteínas y de otras moléculas orgánicas que no pueden escapar de la célula. La mayoría de estas proteínas y otras moléculas orgánicas tienen carga negativa y, por tanto, atraen grandes cantidades de potasio, sodio y también de otros iones positivos. Todas estas moléculas e iones producen ósmosis de agua hacia el interior de la célula. Salvo que este proceso se detenga, la célula se hinchará indefinidamente hasta que explote. El mecanismo normal para impedir este resultado es la bomba Na+K+. la membrana es mucho menos permeable a los iones sodio que a los iones potasio, de modo que una vez que los iones sodio están en el exterior tienen una intensa tendencia a permanecer ahí. Así, este proceso representa una pérdida neta de iones hacia el exterior de la célula, lo que inicia también la ósmosis de agua hacia el exterior de la célula. Si una célula comienza a hincharse por cualquier motivo, la bomba Na+-K+ se activa automáticamente, moviendo aún más iones hacia el exterior y transportando agua con ellos. Por tanto, la bomba Na+-K+ realiza una función continua de vigilancia para mantener el volumen celular normal.
Naturaleza electrógena de la bomba Na+-K+
El hecho de que la bomba Na+-K+ desplace tres iones Na+ hacia el exterior por cada dos iones K+ que desplaza hacia el interior significa que se desplaza una carga positiva neta desde el interior de la célula hasta el exterior en cada ciclo de bombeo. Esta acción genera positividad en el exterior de la célula, aunque produce un déficit de iones positivos en el interior de la célula; es decir, produce negatividad en el interior. Por tanto, se dice que la bomba Na+-K+ es electrógena porque genera un potencial eléctrico a través de la membrana celular.
Transporte activo primario de iones calcio
Los iones calcio normalmente se mantienen a una concentración muy baja en el citosol intracelular de prácticamente todas las células del cuerpo, a una concentración aproximadamente 10.000 veces menor que en el líquido extracelular. Este nivel de mantenimiento se consigue principalmente mediante dos bombas de calcio que funcionan mediante transporte activo primario. Una de ellas, que está en la membrana celular, bombea calcio hacia el exterior de la célula. La otra bombea iones calcio hacia uno o más de los orgánulos vesiculares intracelulares de la célula, como el retículo sarcoplásmico de las células musculares y las mitocondrias en todas las células. En todos estos casos la proteína transportadora penetra en la membrana y actúa como una enzima ATPasa, con la misma capacidad de escindir el ATP que la ATPasa de la proteína transportadora de sodio. La diferencia es que esta proteína tiene un punto de unión muy específico para el calcio en lugar de para el sodio.
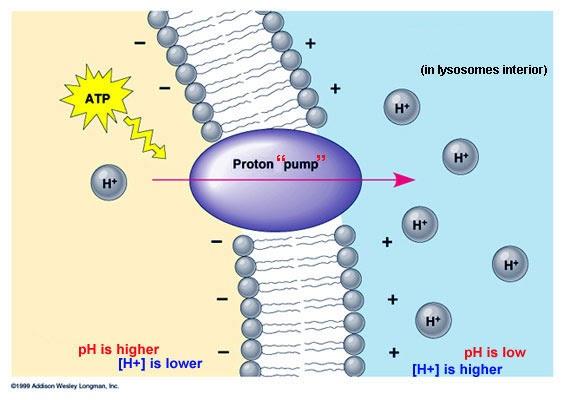
Transporte activo primario de iones hidrógeno
Este transporte activo primario es importante en dos localizaciones del cuerpo:
1) en las glándulas gástricas del estómago: las células parietales que están en las capas profundas tienen el mecanismo activo primario más potente de transporte de iones hidrógeno de todo el cuerpo. Este mecanismo es la base para secretar ácido clorhídrico en las secreciones digestivas del estómago. En el extremo secretor de las células parietales de las glándulas gástricas la concentración del ion hidrógeno aumenta hasta un millón de veces y después se libera hacia el estómago junto con iones cloruro para formar ácido clorhídrico.
2) en la porción distal de los túbulos distales y en los conductos colectores corticales de los riñones: en este caso se secretan grandes cantidades de iones hidrógeno desde la sangre hacia la orina con el objetivo de eliminar de los líquidos corporales el exceso de iones hidrógeno. Los iones hidrógeno se pueden segregar hacia la orina contra un gradiente de concentración de aproximadamente 900 veces.
Transporte activo secundario: cotransporte y contratransporte
Cuando los iones sodio se transportan hacia el exterior de las células mediante transporte activo primario habitualmente se establece un gran gradiente de concentración de iones sodio a través de la membrana celular, con una concentración elevada fuera de la célula y una concentración baja en su interior. Este gradiente representa un almacén de energía porque el exceso de sodio en el exterior de la membrana celular siempre intenta difundir hacia el interior. En condiciones adecuadas esta energía de difusión del sodio puede arrastrar otras sustancias junto con el sodio a través de la membrana celular. Este fenómeno, denominado cotransporte, es una forma de transporte activo secundario. Para que el sodio arrastre otra sustancia con él es necesario un mecanismo de acoplamiento, lo cual se consigue por medio de otra proteína transportadora de la membrana celular. En este caso el transportador actúa como punto de unión tanto para el ion sodio como para la sustancia que se va a cotransportar. Una vez que los dos están unidos, el gradiente de energía del ion sodio hace que este ion y la otra sustancia sean transportados juntos hacia el interior de la célula.
En el contratransporte, los iones sodio intentan una vez más difundir hacia el interior de la célula debido a su gran gradiente de concentración. Sin embargo, esta vez la sustancia que se va a transportar está en el interior de la célula y se debe transportar hacia el exterior. Por tanto, el ion sodio se une a la proteína transportadora en el punto en el que se proyecta hacia la superficie exterior de la membrana, mientras que la sustancia que se va a contratransportar se une a la proyección interior de la proteína transportadora. Una vez que ambos se han unido se produce un cambio conformacional y la energía liberada por la acción del ion sodio que se mueve hacia el interior hace que la otra sustancia se mueva hacia el exterior.
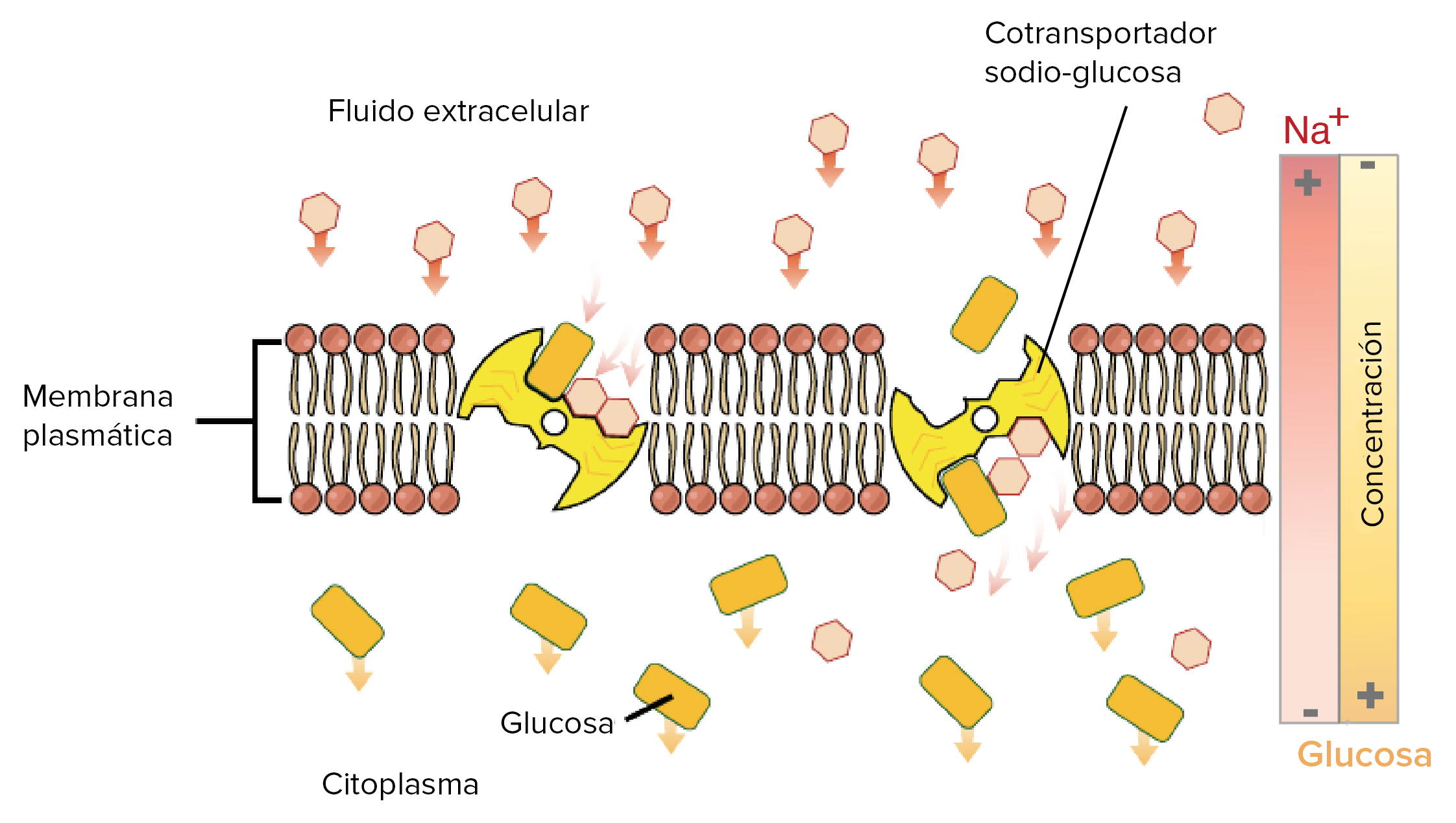
Cotransporte de glucosa y aminoácidos junto con iones sodio
La glucosa y muchos aminoácidos se transportan hacia el interior de la mayor parte de las células contra grandes gradientes de concentración; el mecanismo de esta acción es totalmente mediante cotransporte. Se debe observar que la proteína transportadora tiene dos puntos de unión en su cara externa, uno para el sodio y otro para la glucosa. Además, la concentración de los iones sodio es alta en el exterior y baja en el interior, lo que suministra la energía para el transporte. Una propiedad especial de la proteína transportadora es que no se producirá un cambio conformacional que permita el movimiento de sodio hacia el interior hasta que también una molécula de glucosa se una. Cuando ambos están unidos se produce el cambio conformacional y el sodio y la glucosa son transportados al mismo tiempo hacia el interior de la célula. Por tanto, este es un mecanismo de cotransporte sodio-glucosa. Los cotransportadores de sodio-glucosa son mecanismos especialmente importantes en el transporte de la glucosa a través de las células epiteliales renales e intestinales.
El cotransporte con sodio de los aminoácidos se produce de la misma manera que para la glucosa, excepto porque utiliza un grupo diferente de proteínas transportadoras. Se han identificado al menos cinco proteínas transportadoras de aminoácidos, cada una de las cuales es responsable de transportar un grupo de aminoácidos con características moleculares específicas. El cotransporte con sodio de la glucosa y de los aminoácidos se produce especialmente a través de las células epiteliales del tubo digestivo y de los túbulos renales para favorecer la absorción de estas sustancias hacia la sangre. Este proceso se analizará en capítulos posteriores. Otros mecanismos importantes de cotransporte al menos en algunas células incluyen cotransporte de iones cloruro, yoduro, hierro y urato.
Contratransporte con sodio de iones calcio e hidrógeno
Dos mecanismos de contratransporte especialmente importantes son el contratransporte sodio-calcio y el contratransporte sodio-hidrógeno. El contratransporte sodio-calcio se produce a través de todas o casi todas las membranas celulares, de modo que los iones sodio se mueven hacia el interior y los iones calcio hacia el exterior, ambos unidos a la misma proteína transportadora en un modo de contratransporte. Este mecanismo se produce además del transporte activo primario de calcio que se produce en algunas células. El contratransporte sodio-hidrógeno se produce en varios tejidos. Un ejemplo especialmente importante se produce en los túbulos proximales de los riñones, en los que los iones sodio se desplazan desde la luz del túbulo hacia el interior de la célula tubular, mientras que los iones hidrógeno son contratransportados hacia la luz tubular.
Transporte activo a través de capas celulares
En muchas localizaciones del cuerpo se deben transportar sustancias a través de todo el espesor de una capa celular en lugar de simplemente a través de la membrana celular. El transporte de este tipo se produce a través de:
1) el epitelio intestinal
2) el epitelio de los túbulos renales
3) el epitelio de todas las glándulas exocrinas
4) el epitelio de la vesícula biliar
5) la membrana del plexo coroideo del cerebro, junto con otras membranas.
El mecanismo básico para el transporte de una sustancia a través de una lámina celular es:
1)transporte activo a través de la membrana celular de un polo de las células transportadoras de la capa
2) difusión simple o difusión facilitada a través de la membrana del polo opuesto de la célula.
Hay potenciales eléctricos a través de las membranas de todas las células del cuerpo. Algunas células, como las células nerviosas y musculares, generan impulsos electroquímicos rápidamente cambiantes en sus membranas, y estos impulsos se utilizan para transmitir señales a través de las membranas de los nervios y de los músculos. En otros tipos de células, como las células glandulares, los macrófagos y las células ciliadas, los cambios locales de los potenciales de membrana también activan muchas de las funciones de las células
Física básica de los potenciales de membrana
Potenciales de membrana provocados por concentración de iones
Diferencias a través de una membrana permeable selectiva
la concentración de potasio es grande dentro de la membrana de una fibra nerviosa, pero muy baja fuera de esta. Imaginemos en este caso que la membrana es permeable a los iones potasio, pero no a ningún otro ion. Debido al gran gradiente de concentración de potasio desde el interior hacia el exterior hay una intensa tendencia a que cantidades adicionales de iones potasio difundan hacia fuera a través de la membrana. A medida que lo hacen transportan cargas eléctricas positivas hacia el exterior, generando de esta manera electropositividad fuera de la membrana y electronegatividad en el interior debido a los aniones negativos que permanecen detrás y que no difunden hacia fuera con el potasio. En un plazo de aproximadamente 1 ms la diferencia de potencial entre el interior y el exterior, denominada potencial de difusión, se hace lo suficientemente grande como para bloquear la difusión adicional neta de potasio hacia el exterior, a pesar del elevado gradiente de concentración iónica de potasio. En la fibra nerviosa normal del mamífero la diferencia de potencial es de aproximadamente 94 mV, con negatividad en el interior de la membrana de la fibra.
La ecuación de Nernst describe la relación del potencial de difusión con la diferencia de concentración de iones a través de una membrana
El nivel del potencial de difusión a través de una membrana que se opone exactamente a la difusión neta de un ion particular a través de la membrana se denomina potencial de Nernst para ese ion. La magnitud del potencial de Nernst viene determinada por el cociente de las concentraciones de ese ion específico en los dos lados de la membrana. Cuanto mayor es este cociente, mayor es la tendencia del ion a difundir en una dirección y, por tanto, mayor será el potencial de Nernst necesario para impedir la difusión neta adicional. Se puede utilizar la siguiente ecuación, denominada ecuación de Nernst, para calcular el potencial de Nernst para cualquier ion univalente a la temperatura corporal normal (37 °C):
donde FEM es la fuerza electromotriz y z es la carga eléctrica del ion (p. ej., +1 para K+).
La ecuación de Goldman se utiliza para calcular el potencial de difusión cuando la membrana es permeable a varios iones diferentes
Cuando una membrana es permeable a varios iones diferentes, el potencial de difusión que se genera depende de tres factores:
1) la polaridad de la carga eléctrica de cada uno de los iones.
2) la permeabilidad de la membrana (P) a cada uno de los iones.
3) las concentraciones (C) de los respectivos iones en el interior (i) y en el exterior (e) de la membrana.
Así, la fórmula siguiente, que se denomina ecuación de Goldman o ecuación de Goldman-Hodgkin-Katz, da el potencial de membrana calculado en el interior de la membrana cuando participan dos iones positivos univalentes, sodio (Na+) y potasio (K+), y un ion negativo univalente, cloruro (Cl–).
A partir de la ecuación de Goldman se hacen evidentes varios puntos clave. En primer lugar, los iones sodio, potasio y cloruro son los iones más importantes que participan en la generación de los potenciales de membrana en las fibras nerviosas y musculares, así como en las células neuronales del sistema nervioso. Segundo, el grado de importancia de cada uno de los iones en la determinación del voltaje es proporcional a la permeabilidad de la membrana para ese ion particular. Tercero, un gradiente positivo de concentración iónica desde el interior de la membrana hacia el exterior produce electronegatividad en el interior de la membrana. Cuarto, la permeabilidad de los canales de sodio y de potasio experimenta cambios rápidos durante la transmisión de un impulso nervioso, mientras que la permeabilidad de los canales de cloruro no se modifica mucho durante este proceso. Por tanto, los cambios rápidos de la permeabilidad al sodio y el potasio son los principales responsables de la transmisión de las señales en las neuronas.
Potencial de membrana en reposo de las neuronas
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV. Es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de esta.
Transporte activo de los iones sodio y potasio a través de la membrana: la bomba sodio-potasio (Na+-K+)
todas las membranas celulares del cuerpo tienen una potente bomba Na+-K+ que transporta continuamente iones sodio hacia el exterior de la célula e iones potasio hacia el interior. se trata de una bomba electrógena porque se bombean más cargas positivas hacia el exterior que hacia el interior (tres iones Na+ hacia el exterior por cada dos iones K+ hacia el interior), dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular.
Fuga de potasio a través de la membrana celular nerviosa
un canal proteico, a veces denominado canal de potasio de dominios de poro en tándem, o canal de «fuga» de potasio (K+), en la membrana nerviosa a través de la que pueden escapar iones potasio incluso en una célula en reposo. los canales de fuga de K+ también pueden dejar que se pierdan algunos iones sodio pero los canales son mucho más permeables al potasio que al sodio, en general aproximadamente 100 veces más permeables.
Potencial de acción de las neuronas
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega a su extremo.
Las sucesivas fases del potencial de acción son las siguientes.
Fase de reposo
La fase de reposo es el potencial de membrana en reposo antes del comienzo del potencial de acción. Se dice que la membrana está «polarizada» durante esta fase debido al potencial de membrana negativo de – 90 mV que está presente.
Fase de despolarización
En este momento la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado «polarizado» normal de –90 mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva, un proceso denominado despolarización. En las fibras nerviosas grandes el gran exceso de iones sodio positivos que se mueven hacia el interior hace que el potencial de membrana realmente se «sobreexcite» más allá del nivel cero y que se haga algo positivo. En algunas fibras más pequeñas, así como en muchas neuronas del sistema nervioso central, el potencial simplemente se acerca al nivel cero y no hay sobreexcitación hacia el estado positivo.
Fase de repolarización
En un plazo de algunas diezmilésimas de segundo después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal, que se denomina repolarización de la membrana.
potencial de acción
Canales de sodio y potasio activados por el voltaje
El actor necesario en la producción tanto de la despolarización como de la repolarización de la membrana nerviosa durante el potencial de acción es el canal de sodio activado por el voltaje. Un canal de potasio activado por el voltaje también tiene una función importante en el aumento de la rapidez de la repolarización de la membrana. Estos dos canales activados por el voltaje tienen una función adicional a la de la bomba Na+-K+ y de los canales de fuga de K+.
Activación e inactivación del canal de sodio activado por el voltaje
Este canal tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación. en reposo normal, cuando el potencial de membrana es de –90 mV, la compuerta de activación está cerrada, lo que impide la entrada de iones sodio hacia el interior de la fibra a través de estos canales de sodio.
Activación del canal de sodio
Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje (habitualmente algún punto entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta, que bascula totalmente hasta la posición de abierta. Durante este estado activado, los iones sodio pueden atravesar el canal, aumentando la permeabilidad de la membrana al sodio hasta 500 a 5.000 veces.
Inactivación del canal de sodio
El mismo aumento de voltaje que abre la compuerta de activación también cierra la compuerta de inactivación. Sin embargo, la compuerta de inactivación se cierra algunas diezmilésimas de segundo después de que se abra la compuerta de activación. Es decir, el cambio conformacional que hace bascular la compuerta de inactivación hacia el estado cerrado es un proceso algo más lento que el cambio conformacional que abre la compuerta de activación. Por tanto, después de que el canal de sodio haya permanecido abierto durante algunas diezmilésimas de segundo se cierra la compuerta de inactivación y los iones sodio ya no pueden pasar hacia el interior de la membrana. En este punto el potencial de membrana comienza a recuperarse de nuevo hacia el estado de membrana en reposo, lo que es el proceso de repolarización.
Canal de potasio activado por el voltaje y su activación
el canal de potasio activado por el voltaje en dos estados: durante el estado de reposo y hacia el final del potencial de acción. Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90 mV hacia cero, este voltaje produce una apertura conformacional de la compuerta y permite el aumento de la difusión de potasio hacia fuera a través del canal. Sin embargo, debido a la ligera demora de la apertura de los canales de potasio, en su mayor parte, se abren al mismo tiempo que están comenzando a cerrarse los canales de sodio debido a su inactivación. Por tanto, la disminución de la entrada de sodio hacia la célula y el aumento simultáneo de la salida de potasio desde la célula se combinan para acelerar el proceso de repolarización, lo que da lugar a la recuperación completa del potencial de membrana en reposo en otras pocas diezmilésimas de segundo.
Funciones de otros iones durante el potencial de acción
Se deben considerar al menos otros dos tipos de iones: los aniones negativos y los iones calcio.
Iones con carga negativa (aniones) no difusibles en el interior del axón nervioso
En el interior del axón hay muchos iones de carga negativa que no pueden atravesar los canales de la membrana. Incluyen los aniones de las moléculas proteicas y de muchos compuestos de fosfato orgánicos, compuestos de sulfato y similares. Como estos iones no pueden salir del interior del axón, cualquier déficit de iones positivos en el interior de la membrana deja un exceso de estos aniones negativos no difusibles. Por tanto, estos iones negativos no difusibles son responsables de la carga negativa en el interior de la fibra cuando hay un déficit neto de iones potasio de carga positiva y de otros iones positivos.
Iones calcio
Las membranas de casi todas las células del cuerpo tienen una bomba de calcio similar a la bomba de sodio, y el calcio coopera con el sodio (o actúa en su lugar) en algunas células para producir la mayor parte del potencial de acción. Al igual que la bomba de sodio, la bomba de potasio transporta iones calcio desde el interior hacia el exterior de la membrana celular (o hacia el interior del retículo endoplásmico de la célula), creando un gradiente de ion calcio de aproximadamente 10.000 veces. Además, hay canales de calcio activados por el voltaje. Dado que la concentración de iones calcio es más de 10.000 veces mayor en el fluido extracelular que en el intracelular, existe un enorme gradiente de difusión para el flujo pasivo de iones calcio a las células. Estos canales son ligeramente permeables a los iones sodio y a los iones calcio; sin embargo, su permeabilidad al calcio es aproximadamente 1.000 veces mayor que al sodio en condiciones fisiológicas normales. Cuando el canal se abre como respuesta a un estímulo que despolariza la membrana celular, los iones calcio fluyen al interior de la célula. Una función importante de los canales de iones calcio activados por el voltaje consiste en su contribución a la fase de despolarización en el potencial de acción en algunas células. No obstante, la activación de los canales de calcio es lenta, y precisa hasta 10 a 20 veces más tiempo para su activación que los canales de sodio. Por este motivo, a menudo se denominan canales lentos, en contraposición a los canales de sodio, que se denominan canales rápidos. Por tanto, la apertura de los canales de calcio proporciona una despolarización más sostenida, mientras que los canales de sodio desempeñan un papel clave en la iniciación de los potenciales de acción. Hay abundantes canales de calcio tanto en el músculo cardíaco como el músculo liso.
Aumento de la permeabilidad de los canales de sodio cuando hay déficit de iones calcio
La concentración de iones calcio en el líquido extracelular también tiene un efecto profundo sobre el nivel de voltaje al que se activan los canales de sodio. Cuando hay déficit de iones calcio, los canales de sodio se activan (abren) por un pequeño aumento del potencial de membrana desde su nivel normal, muy negativo. Por tanto, la fibra nerviosa se hace muy excitable, y a veces descarga de manera repetitiva sin provocación en lugar de permanecer en su estado de reposo. De hecho, es necesario que la concentración del ion calcio disminuya solo un 50% por debajo de su concentración normal para que se produzca la descarga espontánea en algunos nervios periféricos, produciendo con frecuencia «tetania» muscular. Esta tetania muscular a veces resulta mortal por la contracción tetánica de los músculos respiratorios.
Inicio del potencial de acción
Un ciclo de retroalimentación positiva abre los canales de sodio
siempre que no haya alteraciones de la membrana de la fibra nerviosa, no se produce ningún potencial de acción en el nervio normal. Sin embargo, si algún episodio produce una elevación suficiente del potencial de membrana desde –90 mV hacia el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos canales de sodio activados por el voltaje. Esta situación permite la entrada rápida de iones sodio, lo que produce una elevación adicional del potencial de membrana y abre aún más canales de sodio activados por el voltaje y permite que se produzca una mayor entrada de iones sodio hacia el interior de la fibra. Este proceso es un círculo vicioso de retroalimentación positiva que, una vez que la retroalimentación es lo suficientemente intensa, continúa hasta que se han activado (abierto) todos los canales de sodio activados por el voltaje. Posteriormente, en un plazo de otra fracción de milisegundo, el aumento del potencial de membrana produce cierre de los canales de sodio y apertura de los canales de potasio, y pronto finaliza el potencial de acción.
Umbral para el inicio del potencial de acción
No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva. Esto se produce cuando el número de iones sodio que entran en la fibra supera al número de iones potasio que salen. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Por tanto, un aumento súbito del potencial de membrana en una fibra nerviosa grande desde –90 mV hasta aproximadamente –65 mV suele dar lugar a la aparición explosiva de un potencial de acción. Se dice que este nivel de –65 mV es el umbral para la estimulación.
Propagación del potencial de acción
el potencial de acción que se produce en un punto de la membrana. Sin embargo, un potencial de acción que se desencadena en cualquier punto de una membrana excitable habitualmente excita porciones adyacentes de la membrana, dando lugar a la propagación del potencial de acción a lo largo de la membrana.
Dirección de la propagación
una membrana excitable no tiene una dirección de propagación única, sino que el potencial de acción viaja en todas las direcciones alejándose del estímulo (incluso a lo largo de todas las ramas de una fibra nerviosa) hasta que se ha despolarizado toda la membrana.
Principio del todo o nada
Una vez que se ha originado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización viaja por toda la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no lo son. Este principio se denomina principio del todo o nada y se aplica a todos los tejidos excitables normales.
Restablecimiento de los gradientes iónicos de sodio y potasio tras completarse los potenciales de acción: la importancia del metabolismo de la energía
La propagación de cada potencial de acción a lo largo de una fibra nerviosa reduce ligeramente las diferencias de concentración de sodio y de potasio en el interior y en el exterior de la membrana, porque los iones sodio difunden hacia el interior durante la despolarización y los iones potasio difunden hacia el exterior durante la repolarización. Para un único potencial de acción este efecto es tan pequeño que no se puede medir. los iones sodio que han difundido hacia el interior de la célula durante los potenciales de acción y los iones potasio que han difundido hacia el exterior deben volver a su estado original por la bomba Na+-K+. Como esta bomba precisa energía para esta operación, esta «recarga» de la fibra nerviosa es un proceso metabólico activo que utiliza la energía que procede del sistema energético del trifosfato de adenosina de la célula.
Una característica especial de la bomba Na+-K+-adenosina trifosfatasa es que su grado de actividad se estimula mucho cuando se acumula un exceso de iones sodio en el interior de la membrana celular. De hecho, la actividad de bombeo aumenta aproximadamente en proporción a la tercera potencia de esta concentración intracelular de sodio. Cuando la concentración interna de sodio aumenta desde 10 hasta 20 mEq/l, la actividad de la bomba no solo aumenta, sino que lo hace aproximadamente ocho veces.
Meseta en algunos potenciales de acción
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y solo después comienza la repolarización. Este tipo de potencial de acción se produce en las fibras musculares cardíacas, en las que la meseta dura hasta 0,2 a 0,3 s y hace que la contracción del músculo cardíaco dure este mismo y prolongado período de tiempo.
La causa de la meseta es una combinación de varios factores. En primer lugar, en el proceso de despolarización del músculo cardíaco participan dos tipos de canales: 1) los canales de sodio habituales activados por el voltaje, denominados canales rápidos, y 2) los canales de calcio-sodio activados por el voltaje (canales de calcio de tipo L), que tienen una apertura lenta y que, por tanto, se denominan canales lentos. La apertura de los canales rápidos origina la porción en espiga del potencial de acción, mientras que la apertura prolongada de los canales lentos de calcio-sodio principalmente permite la entrada de iones calcio en la fibra, lo que es responsable en buena medida de la porción de meseta del potencial de acción.
Ritmicidad de algunos tejidos excitables: descarga repetitiva
Las descargas repetitivas autoinducidas aparecen normalmente en el corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso central. Estas descargas rítmicas producen:
1) el latido rítmico del corazón
2) el peristaltismo rítmico de los intestinos
3) fenómenos neuronales, como el control rítmico de la respiración.
Además, casi todos los demás tejidos excitables pueden descargar de manera repetitiva si se reduce lo suficiente el umbral de estimulación de las células del tejido.
Proceso de reexcitación necesario para la ritmicidad espontánea
Para que se produzca ritmicidad espontánea la membrana, incluso en su estado natural, debe ser lo suficientemente permeable a los iones sodio (o a los iones calcio y sodio a través de los canales lentos de calcio-sodio) como para permitir la despolarización automática de la membrana. el potencial de membrana «en reposo» del centro de control rítmico del corazón es de solo – 60 a –70 mV; este voltaje no es lo suficientemente negativo como para mantener totalmente cerrados los canales de sodio y de calcio. Por tanto, se produce la siguiente secuencia:
1) algunos iones sodio y calcio fluyen hacia el interior
2) esta actividad produce aumento del voltaje de la membrana en dirección positiva, que aumenta más la permeabilidad de la membrana
3) se produce flujo de entrada de aún más iones
4)aumenta más la permeabilidad, de manera progresiva, hasta que se genera un potencial de acción.
Después, al final del potencial de acción se repolariza la membrana. Tras otra demora de milisegundos o segundos la excitabilidad espontánea produce una nueva despolarización y se produce espontáneamente un nuevo potencial de acción. Este ciclo continúa de manera indefinida y produce la excitación rítmica autoinducida del tejido excitable.
Características especiales de la transmisión de señales en los troncos nerviosos
Fibras nerviosas mielinizadas y no mielenilizadas
fibra mielinizada típica: el núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de mielina que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier.
Las células de Schwann depositan la vaina de mielina alrededor del axón de la siguiente manera: en primer lugar, la membrana de una célula de Schwann rodea el axón. Después, la célula de Schwann rota muchas veces alrededor del axón, depositando múltiples capas de membrana de la célula de Schwann que contiene la sustancia lipídica esfingomielina. Esta sustancia es un excelente aislante eléctrico que disminuye el flujo iónico a través de la membrana aproximadamente 5.000 veces. En la unión entre dos células de Schwann sucesivas a lo largo del axón permanece una pequeña zona no aislada de solo 2 a 3 μm de longitud en la que los iones pueden seguir fluyendo con facilidad a través de la membrana del axón entre el líquido extracelular y el líquido intracelular del interior del axón. Esta zona se denomina nódulo de Ranvier
Conducción «saltatoria» en las fibras mielinizadas de un nódulo a otro
Aunque los iones apenas pueden fluir a través de las gruesas vainas de mielina de los nervios mielinizados, sí lo hacen fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen solo en los nódulos. A pesar de todo, los potenciales de acción se conducen desde un nódulo a otro, esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro. Así, el impulso nervioso recorre a saltos la fibra, lo que es el origen del término «saltatoria».
Velocidad de conducción en las fibras nerviosas
La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan solo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (un valor superior a la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
Excitación: el proceso de generación del potencial de acción
cualquier factor que haga que los iones sodio comiencen a difundir hacia el interior a través de la membrana en un número suficiente puede desencadenar la apertura regenerativa automática de los canales de sodio. Esta apertura regenerativa automática se puede deber a un trastorno mecánico de la membrana, a los efectos químicos sobre la membrana o al paso de electricidad a través de la membrana.
«Período refractario» tras un potencial de acción, durante el cual no se puede generar un nuevo estímulo
No se puede producir un nuevo potencial de acción en una fibra excitable mientras la membrana siga despolarizada por el potencial de acción precedente. El motivo de esta restricción es que poco después del inicio del potencial de acción se inactivan los canales de sodio (o los canales de potasio, o ambos), y ninguna magnitud de la señal excitadora que se aplique a estos canales en este momento abrirá las compuertas de inactivación. La única situación que permitirá que se vuelvan a abrir es que el potencial de membrana vuelva al nivel del potencial de membrana en reposo original o cerca de este. Entonces, en otra pequeña fracción de segundo se abren las compuertas de inactivación del canal y se puede iniciar un nuevo potencial de acción. El período durante el cual no se puede generar un segundo potencial de acción, incluso con un estímulo intenso, se denomina período refractario absoluto. Para las fibras nerviosas mielinizadas grandes este período es de aproximadamente 1/2.500 s.
Anestésicos locales
Entre los estabilizadores más importantes están las muchas sustancias que se utilizan en clínica como anestésicos locales, como procaína y tetracaína. La mayoría de estas sustancias actúan directamente sobre las compuertas de activación de los canales de sodio, haciendo que sea mucho más difícil abrir estas compuertas, reduciendo de esta manera la excitabilidad de la membrana. Cuando se ha reducido tanto la excitabilidad que el cociente entre en la intensidad del potencial de acción respecto al umbral de excitabilidad (denominado «factor de seguridad») se reduce por debajo de 1, los impulsos nerviosos no pasan a lo largo de los nervios anestesiados.
Tipos de receptores sensitivos y estímulos que detectan
1) mecanorreceptores, que detectan la compresión mecánica o su estiramiento, o el de los tejidos adyacentes; 2) termorreceptores, que detectan los cambios en la temperatura, donde algunos de los receptores se encargan del frío y otros del calor; 3) nocirreceptores (receptores del dolor), que detectan daños físicos o químicos que se producen en los tejidos; 4) receptores electromagnéticos, que detectan la luz en la retina ocular, y 5) quimiorreceptores, que detectan el gusto en la boca, el olfato en la nariz, la cantidad de oxígeno en la sangre arterial, la osmolalidad de los líquidos corporales, la concentración de dióxido de carbono y otros factores que completen la bioquímica del organismo.
Sensibilidad diferencial de los receptores
cada tipo de receptor resulta muy sensible a una clase de estímulo sensitivo para el que está diseñado y en cambio es casi insensible a otras clases.
Modalidad sensitiva: el principio de la línea marcada
Cada uno de los principales tipos sensitivos que podemos experimentar, dolor, tacto, visión, sonido, etc., se llama modalidad de sensación.cada fascículo nervioso termina en un punto específico del sistema nervioso central y el tipo de sensación vivida cuando se estimula una fibra nerviosa queda determinado por la zona del sistema nervioso a la que conduce esta fibra. Esta especificidad de las fibras nerviosas para transmitir nada más que una modalidad de sensación se llama principio de la línea marcada.
Transducción de estímulos sensitivos en impulsos nerviosos
Cualquiera que sea el tipo de estímulo que les excite, su efecto inmediato consiste en modificar su potencial eléctrico de membrana. Este cambio en el potencial se llama potencial de receptor.
Mecanismos de los potenciales de receptor :Los diversos receptores pueden excitarse siguiendo alguno de los siguientes modos de generar potenciales de receptor: 1) por deformación mecánica del receptor, que estire su membrana y abra los canales iónicos; 2) por la aplicación de un producto químico a la membrana, que también abra los canales iónicos; 3) por un cambio de la temperatura de la membrana, que modifique su permeabilidad, o 4) por los efectos de la radiación electromagnética, como la luz que incide sobre un receptor visual de la retina, al modificar directa o indirectamente las características de la membrana del receptor y permitir el flujo de iones a través de sus canales.
Adaptación de los receptores
Otra característica que comparten todos los receptores sensitivos es su adaptación parcial o total a cualquier estímulo constante después de haber transcurrido un tiempo. Es decir, cuando se aplica un estímulo sensitivo continuo, el receptor responde al principio con una frecuencia de impulsos alta y después baja cada vez más hasta que acaba disminuyendo la frecuencia de los potenciales de acción para pasar a ser muy pocos o muchas veces desaparecer del todo.
Los receptores de adaptación lenta detectan la intensidad continua del estímulo: los receptores tónicos
Los receptores de adaptación lenta siguen transmitiendo impulsos hacia el cerebro mientras siga presente el estímulo (o al menos durante muchos minutos u horas). Por tanto, mantienen al cerebro constantemente informado sobre la situación del cuerpo y su relación con el medio. Debido a su capacidad para seguir transmitiendo información durante muchas horas, o incluso días, también se les denomina receptores tónicos.
Los receptores de adaptación rápida detectan cambios en la intensidad del estímulo: «receptores de velocidad, receptores de movimiento o receptores fásicos
Los receptores que se adaptan con rapidez no pueden utilizarse para transmitir una señal continua debido a que solo se activan cuando cambia la intensidad del estímulo. Con todo, reaccionan potentemente siempre que esté teniendo lugar un cambio de hecho
Transmisión de señales de diferente intensidad por los fascículos nerviosos: sumación espacial y temporal
Una de las características de toda señal que siempre ha de transportarse es su intensidad. Los diversos grados de esta variable pueden transmitirse mediante un número creciente de fibras paralelas o enviando más potenciales de acción a lo largo de una sola fibra. Estos dos mecanismos se llaman, respectivamente, sumación espacial y sumación temporal.
Sumación espacial:
el fenómeno de la sumación espacial, por el cual se transmite la intensidad creciente de una señal mediante un número progresivamente mayor de fibras. Así pues, las señales más intensas cada vez se diseminan a más fibras. Este proceso es el fenómeno de la sumación espacial.
Sumación temporal:
Un segundo medio para transmitir señales de intensidad creciente consiste en acelerar la frecuencia de los impulsos nerviosos que recorren cada fibra, lo que se denomina sumación temporal
Transmisión y procesamiento de las señales en grupos neuronales
La zona neuronal estimulada por cada fibra nerviosa que entra se llama campo de estimulación. la neurona más próxima a su «campo» recibe un gran número de terminales derivado de la fibra que entra, pero que esta cantidad es cada vez menor en las neuronas más alejadas
Divergencia de las señales que atraviesan los grupos neuronales
Muchas veces es importante que las señales débiles que penetran en un grupo neuronal acaben excitando a una cantidad mucho mayor de las fibras nerviosas que lo abandonan. Este fenómeno se llama divergencia.Esta divergencia significa sencillamente que una señal de entrada se disemina sobre un número creciente de neuronas a medida que atraviesa sucesivos órdenes de células en su camino.
Convergencia de señales
La convergencia significa que un conjunto de señales procedentes de múltiples orígenes se reúnen para excitar una neurona concreta.la convergencia desde una sola fuente, es decir, numerosos terminales derivados de la llegada de un solo fascículo de fibras acaban en la misma neurona.los potenciales de acción que convergen sobre la neurona desde muchos terminales proporcionan una sumación espacial suficiente para llevar a la célula hasta el umbral necesario de descarga.
Prolongación de una señal por un grupo neuronal: posdescarga
una señal que penetra en un grupo suscita una descarga de salida prolongada, llamada posdescarga, cuya duración va desde unos pocos milisegundos hasta muchos minutos después de que haya acabado la señal de entrada
Circuito reverberante (oscilatorio) como causa de la prolongación de la señal
Uno de los circuitos más importantes del sistema nervioso es el circuito reverberante u oscilatorio. Está ocasionado por una retroalimentación positiva dentro del circuito neuronal que ejerce una retroalimentación encargada de reexcitar la entrada del mismo circuito.
Emisión de señales rítmicas
Muchos circuitos neuronales emiten señales de salida rítmicas: por ejemplo, una señal respiratoria rítmica nace en los centros respiratorios del bulbo raquídeo y de la protuberancia. La señal rítmica respiratoria se mantiene de por vida. Otras señales rítmicas, como las que causan los movimientos de rascado de un perro con la pata trasera o las actividades motoras de cualquier animal al caminar, requieren la existencia de estímulos de entrada en los respectivos circuitos para desencadenar las señales rítmicas.
Fatiga sináptica como medio para estabilizar el sistema nervioso
La fatiga sináptica significa meramente que la transmisión sináptica se vuelve cada vez más débil cuanto más largo e intenso sea el período de excitación
Corrección automática a corto plazo de la sensibilidad de la vía mediante el mecanismo de la fatiga
Cuando están sometidas a un uso excesivo, suelen acabar fatigándose, por lo que desciende su sensibilidad. A la inversa, las que están infrautilizadas se encuentran descansadas y sus sensibilidades aumentan. Por tanto, la fatiga y su recuperación constituyen un medio importante a corto plazo para moderar la sensibilidad de los diferentes circuitos del sistema nervioso.
Cambios a largo plazo en la sensibilidad sináptica ocasionados por la regulación al alza o a la baja de los receptores sinápticos
La sensibilidad a largo plazo de las sinapsis puede cambiar tremendamente si la cantidad de proteínas receptoras presentes en los puntos sinápticos se regula al alza en una situación de baja actividad, y a la baja cuando haya una hiperactividad.
si la sensibilidad de unos cuantos de estos circuitos fuera anormalmente alta; entonces cabría esperar la presencia casi continua de calambres musculares, convulsiones, alteraciones psicóticas, alucinaciones, tensión mental u otros trastornos nerviosos. Afortunadamente,, los controles automáticos suelen reajustar la sensibilidad de los circuitos de vuelta dentro de unos límites de reactividad controlables en cualquier momento en que empiecen a estar demasiado activos o demasiado deprimidos.