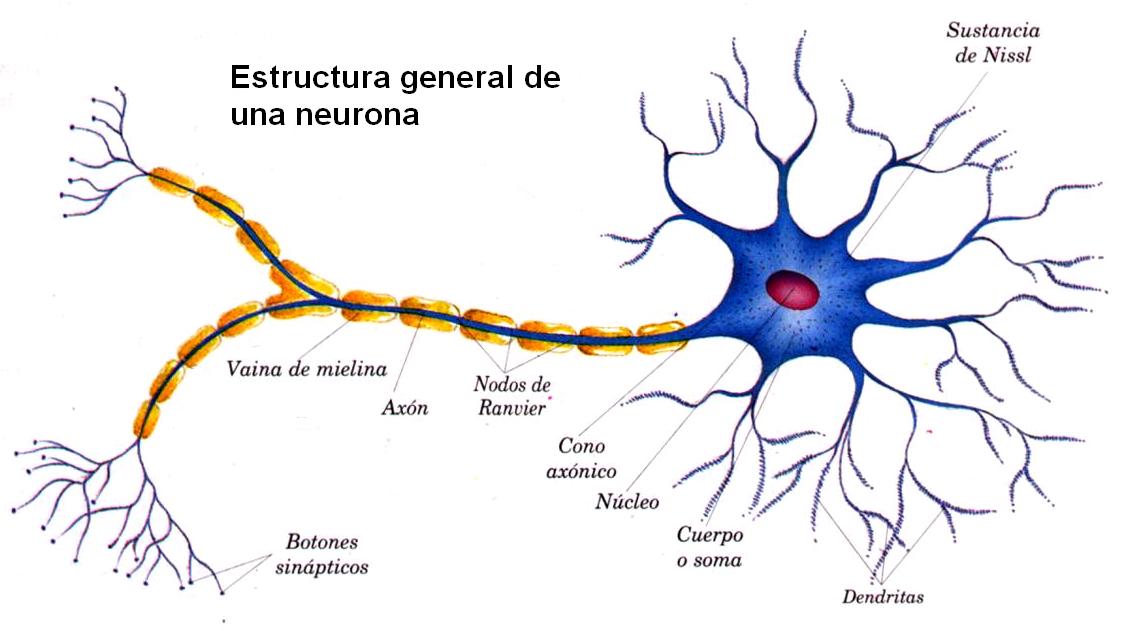
Todos nosotros somos conscientes de los múltiples estados posibles que presenta la actividad cerebral, como el sueño, la vigilia, la excitación extrema, e incluso los diversos estados de ánimo de una persona, entre ellos la euforia, la depresión y el miedo. Cualquiera de estos estados obedece a distintas fuerzas activadoras o inhibidoras generadas normalmente en el encéfalo.
SUEÑO
El sueño se define como el estado de inconsciencia del que puede ser despertada una persona mediante estímulos sensitivos o de otro tipo. Hay que distinguirlo del coma, que es el estado de inconsciencia del que no puede despertarse a una persona. El sueño está integrado por múltiples fases, desde el más ligero hasta el más profundo. Los investigadores que se dedican a este tema también lo dividen en dos tipos totalmente diferentes cuyas cualidades son distintas.

SUEÑO REM ( PARADOJICO )
En una noche normal de sueño, los episodios de sueño REM que duran de 5 a 30 min suelen aparecer en promedio cada 90 min en los adultos jóvenes. Cuando una persona está muy somnolienta, cada episodio de sueño REM es breve e incluso puede estar ausente. Cuando la persona se encuentra más descansada, las duraciones de los episodios REM aumentan. El sueño REM presenta varias características importantes:
- Es una forma activa de sueño que se asocia comúnmente con los sueños y con movimientos activos de los músculos del cuerpo.
- Resulta más difícil despertar a la persona con estímulos sensoriales que durante el sueño de ondas lentas, y habitualmente las personas se despiertan de forma espontánea por la mañana durante un episodio de sueño REM.
- El tono muscular en todo el cuerpo se encuentra muy deprimido, lo que indica una fuerte inhibición de las zonas espinales de control de los músculos.
- Las frecuencias cardíaca y respiratoria suelen volverse irregulares, un hecho característico del estado de sueño.
- A pesar de la extrema inhibición de los músculos periféricos, se producen movimientos musculares irregulares además de los movimientos rápidos de los ojos.
- El encéfalo está muy activo en el sueño REM, y el metabolismo encefálico general puede incrementarse hasta en un 20%
SUEÑO DE ONDAS LENTAS ( NO REM )
Este sueño resulta sumamente reparador y va asociado a un descenso del tono vascular periférico y de otras muchas funciones
vegetativas del cuerpo. Por ejemplo, se produce una disminución del 10 al 30% en la presión arterial, la frecuencia respiratoria y el índice metabólico basal. Aunque al sueño de ondas lentas se le llama a menudo «sueño sin sueños», durante su transcurso hay sueños y, en ocasiones, hasta pesadillas. La diferencia entre los sueños presentes en el sueño de ondas lentas y los que suceden en el sueño REM estriba en que estos últimos van asociados a una mayor actividad muscular del cuerpo. Además, los del sueño de ondas lentas no suelen recordarse porque no tiene lugar la consolidación de los sueños en la memoria.

Centros nerviosos, sustancias neurohumorales y mecanismos capaces de causar sueño: posible función específica de la serotonina
La estimulación de diversas zonas específicas del encéfalo puede producir un sueño dotado de unas
características próximas a las del sueño natural. Entre ellas figuran las siguientes:
- La zona de estimulación para generar un sueño casi natural más constante son los núcleos del rafe en la mitad inferior de la protuberancia y en el bulbo raquídeo. Estos núcleos comprenden una lámina fina de neuronas especiales situadas en la línea media. Las fibras nerviosas que nacen en ellos se diseminan a nivel local por la formación reticular del tronco del encéfalo y también ascienden hacia el tálamo, el hipotálamo, la mayor parte de las regiones del sistema límbico e incluso hasta la neocorteza cerebral. Además, otras fibras descienden hacia la médula espinal, y acaban en las astas posteriores, donde son capaces de inhibir las señales sensitivas recibidas, incluido el dolor. Muchas terminaciones nerviosas de las fibras procedentes de estas neuronas del rafe segregan serotonina. Si a un animal se le administra un fármaco que bloquee su formación, muchas veces no puede dormir a lo largo de varios días después. Por tanto, se ha supuesto que la serotonina es una sustancia transmisora vinculada a la producción del sueño.


CICLO DEL SUEÑO-VIGILIA

Cuando los centros del sueño no están activos, los núcleos reticulares activadores del mesencéfalo y la parte superior de la protuberancia se encuentran liberados de su inhibición, lo que les permite una activación espontánea. Esta actividad espontánea a su vez excita a la corteza cerebral y al sistema nervioso periférico, los cuales devuelven numerosas señales de retroalimentación positiva a los mismos núcleos reticulares activadores para estimularles aún más. Por tanto, una vez que comienza la vigilia, su tendencia natural la lleva a mantenerse por sí sola debido a toda esta actividad de retroalimentación positiva. A continuación, después de que el encéfalo haya permanecido activo muchas horas, se supone que hasta las neuronas del sistema activador acaban por fatigarse. Por consiguiente, el ciclo de retroalimentación positiva entre los núcleos reticulares mesencefálicos y la corteza cerebral decae, y se ve relevado por los efectos hipnóticos a cargo de los centros del sueño, lo que da lugar a una veloz transición de nuevo hasta dicho estado desde la vigilia.

LAS NEURONAS OREXIGENAS SON IMPORTANTES EN EL DESPERTAR Y LA VIGILA
La orexina (también denominada hipocretina) es producida en el hipotálamo por neuronas que proporcionan estímulos aferentes excitadores a muchas otras áreas del encéfalo en las que existen receptores de orexina. Las neuronas orexígenas están más activas durante la vigilia y casi dejan de activarse durante el sueño de ondas lentas y el sueño REM. La pérdida de señales orexígenas a consecuencia de la presencia de receptores anómalos de orexina o de la destrucción de neuronas productoras de orexina provoca narcolepsia, un trastorno del sueño caracterizado por una somnolencia excesiva durante el día y ataques súbitos de sueño que pueden producirse incluso mientras la persona afectada está hablando o trabajando

FUNCIONES FISIOLÓGICAS DEL SUEÑO
una ligera restricción del sueño durante unos días puede deteriorar el rendimiento cognitivo y físico, la productividad general y la salud de una persona.La vigilia prolongada suele asociarse a una disfunción progresiva de los procesos mentales y en ocasiones da lugar incluso a comportamientos anormales. Todos estamos familiarizados con la mayor torpeza de pensamiento que aparece hacia el final de un período de vigilia prolongado, pero, además, una persona puede volverse irritable o incluso adquirir rasgos psicóticos después de verse forzada a mantener este estado. Por tanto, podemos suponer que el sueño restablece por múltiples vías los niveles oportunos de actividad cerebral y el «equilibrio» normal entre las diversas funciones del sistema nervioso central. Se ha postulado que el sueño sirve para muchas funciones, como son: 1) la madurez nerviosa; 2) la facilitación del aprendizaje o la memoria; 3) la cognición; 4) la eliminación de los productosbmetabólicos de desecho generados por la actividad nerviosa en el encéfalo despierto, y 5) la conservación de energía metabólica

ONDAS CEREBRALES
Los registros eléctricos recogidos en la superficie cerebral o incluso en la superficie de la cabeza ponen de manifiesto que existe una actividad eléctrica constante en el encéfalo. Tanto la intensidad como los patrones de esta variable vienen determinados por el grado de excitación que presentan sus diversos componentes como consecuencia del sueño, la vigilia o trastornos cerebrales como la epilepsia o incluso las psicosis. Las ondulaciones de los potenciales eléctricos recogidos, se llaman ondas cerebrales, y el registro en su integridad recibe el nombre de electroencefalograma (EEG).

En las personas sanas, la mayoría de las ondas del EEG pueden clasificarse como ondas α, β, θ y δ.
Las ondas α son ondas rítmicas, con una frecuencia entre 8 y 13 ciclos/s, y que están presentes en el EEG de casi todos los adultos sanos mientras permanecen despiertos y en un estado de reposo tranquilo en su actividad cerebral. Estas ondas adquieren mayor intensidad en la región occipital, pero también pueden recogerse en las regiones parietal y frontal del cuero cabelludo. Su voltaje suele ser de unos 50 μV. Las ondas α desaparecen durante el sueño profundo.

Las ondas β presentan unas frecuencias superiores a 14 ciclos/s y llegan hasta los 80. Se registran sobre todo en las regiones parietal y frontal durante la activación específica de estas partes del cerebro.
Las ondas θ tienen unas frecuencias entre 4 y 7 ciclos/s. Aparecen normalmente en los niños en las regiones parietal y temporal, pero también en algunos adultos ante situaciones de estrés emocional, especialmente en circunstancias de desánimo y de frustración. Asimismo, las ondas θ están presentes en muchos trastornos nerviosos, con frecuencia en los estados degenerativos cerebrales.
Las ondas δ engloban todas las ondas del EEG con frecuencias menores a 3,5 ciclos/s, y a menudo poseen voltajes del doble al cuádruple que la mayoría de los demás tipos de ondas cerebrales. Se dan a lo largo del sueño muy profundo, en la lactancia y en personas con enfermedades orgánicas serias del cerebro
CONVULSIONES Y EPILEPSIA
Las convulsiones son interrupciones temporales de la función encefálica causadas por una actividad neuronal excesiva e incontrolada. Según la distribución de las descargas neuronales, las manifestaciones de las convulsiones pueden estar comprendidas entre fenómenos experienciales apenas perceptibles y convulsiones espectaculares. Estas convulsiones sintomáticas temporales no suelen persistir si se corrige el trastorno subyacente. Pueden estar causadas por múltiples dolencias neurológicas o médicas, como trastornos agudos de electrólitos, hipoglucemia, fármacos (p. ej., cocaína), eclampsia, insuficiencia renal, encefalopatía hipertensiva, meningitis, y así sucesivamente.


la epilepsia es una enfermedad crónica de convulsiones recurrentes que también puede oscilar entre síntomas breves y casi indetectables y períodos de vigorosa agitación y convulsiones. La epilepsia no es una enfermedad única. Sus síntomas clínicos son heterogéneos y reflejan múltiples causas subyacentes y mecanismos fisiopatológicos que provocan disfunción cerebral y lesiones, como traumatismos, tumores, infección o cambios degenerativos. Los factores hereditarios parecen ser importantes, aunque en muchos pacientes no es posible identificar una causa específica y pueden coexistir varios factores, lo que refleja un estado patológico adquirido del encéfalo y una predisposición genética.
Las crisis epilépticas pueden clasificarse en dos tipos principales: 1) crisis focales (tambiénndenominadas crisis parciales), que se limitan a un área focal de un hemisferio cerebral, y 2) crisis generalizadas, que afectan de forma difusa a los dos hemisferios de la corteza cerebral. Sin embargo, las crisis parciales a veces pueden evolucionar a formas generalizadas.

Tratamiento de la epilepsia
La mayoría de los fármacos disponibles actualmente para tratar la epilepsia parecen bloquear el inicio o la extensión de las convulsiones, aunque no se conoce la forma exacta de acción de algunos de ellos, o bien tal vez realicen múltiples acciones. Algunos de los principales efectos de los diversos fármacos antiepilépticos son: 1) bloqueo de los canales del sodio dependientes del voltaje 2) alteración de las corrientes de calcio 3) aumento en la actividad GABA 4) inhibición de los receptores de glutamato, el neurotransmisor excitador más común y 5) múltiples mecanismos de acción (p. ej., valproato y topiramato, que bloquean los canales del sodio dependientes del voltaje e incrementan los niveles de GABA en el encéfalo). La elección del fármaco antiepiléptico recomendado por las directrices actuales depende del tipo de epilepsia, la edad del paciente y otros factores, aunque, cuando sea posible, la mejor opción es proceder a la corrección de la causa subyacente de las convulsiones.

COMPORTAMIENTO PSICÓTICO Y DEMENCIA: FUNCIONES DE LOS SISTEMAS NEUROTRANSMISORES ESPECÍFICOS
Los estudios clínicos de pacientes con diversas psicosis o con distintos tipos de demencia han dado a entender que muchos de estos procesos obedecen a un menor funcionamiento de las neuronas que segregan un neurotransmisor específico.
Depresión y psicosis maníaco-depresiva: disminución de la actividad de los sistemas neurotransmisores de noradrenalina y serotonina
Se han acumulado muchas pruebas indicativas de que la depresión mental psicótica, podría estar causada por un descenso de la formación de noradrenalina, de serotonina o de ambas en el encéfalo.



Los pacientes deprimidos sienten síntomas de pena, tristeza, desesperación y amargura. Además, suelen perder el apetito y el deseo sexual y padecen un insomnio grave; muchas veces asociado a este cuadro hay un estado de agitación psicomotora pese a la depresión. Una razón fundamental para pensar que la depresión podría estar ocasionada por un descenso en la actividad de las neuronas secretoras de noradrenalina y de serotonina reside en que los fármacos capaces de bloquear esta secreción, como reserpina, a menudo provocan dicho trastorno. En cambio, el tratamiento con fármacos que aumenten los efectos excitadores de la noradrenalina y la serotonina en las terminaciones nerviosas puede ser eficaz más o menos en el 70% de los pacientes depresivos.
Algunos pacientes con depresión mental alternan entre la depresión y la manía, lo que se denomina trastorno bipolar o psicosis maníaco-depresiva, y unos pocos pacientes exhiben solo manía, sin los episodios depresivos

Esquizofrenia: posible funcionamiento excesivo de parte del sistema dopaminérgico
La esquizofrenia se manifiesta bajo numerosas variedades. Uno de los tipos más frecuentes se observa en la persona que oye voces y tiene delirios, un temor intenso u otras clases de sentimientos sin un origen real. Muchos esquizofrénicos sufren una gran paranoia, con una sensación de persecución a cargo de alguna fuente externa. Pueden presentar un lenguaje incoherente, disociación de ideas y secuencias anormales de pensamiento, y a menudo se encuentran retraídos, a veces adoptando una postura anormal e incluso rigidez.



Se ha propuesto que el exceso de dopamina en personas con esquizofrenia procede de un grupo de neuronas secretoras de esta sustancia cuyos somas celulares están situados en el tegmento ventral del mesencéfalo, en una posición medial y superior a la sustancia negra. Estas neuronas dan origen al denominado sistema dopaminérgico mesolímbico que envía fibras nerviosas y segrega dopamina hacia las porciones mediales y anteriores del sistema límbico, sobre todo hacia el hipocampo, la amígdala, la zona anterior del núcleo caudado y partes de los lóbulos prefrontales. Todos ellos representan potentes centros para el control del comportamiento.
Enfermedad de Alzheimer: placas amiloides y pérdida de memoria
La enfermedad de Alzheimer se define como el envejecimiento prematuro del encéfalo, que suele comenzar al llegar a la mitad de la vida adulta y progresa con rapidez hasta una enorme pérdida de las capacidades mentales, semejante a la que se observa en las personas muy ancianas. Sus rasgos clínicos son los siguientes: 1) una afectación de la memoria de tipo amnésico; 2) un deterioro del lenguaje, y 3) un déficit visoespacial. Las alteraciones motoras y sensitivas, los trastornos de la marcha y las convulsiones son infrecuentes hasta las últimas fases de la enfermedad. Una observación constante en la enfermedad de Alzheimer es la desaparición neuronal en el componente de la vía límbica que se encargandel proceso de la memoria.