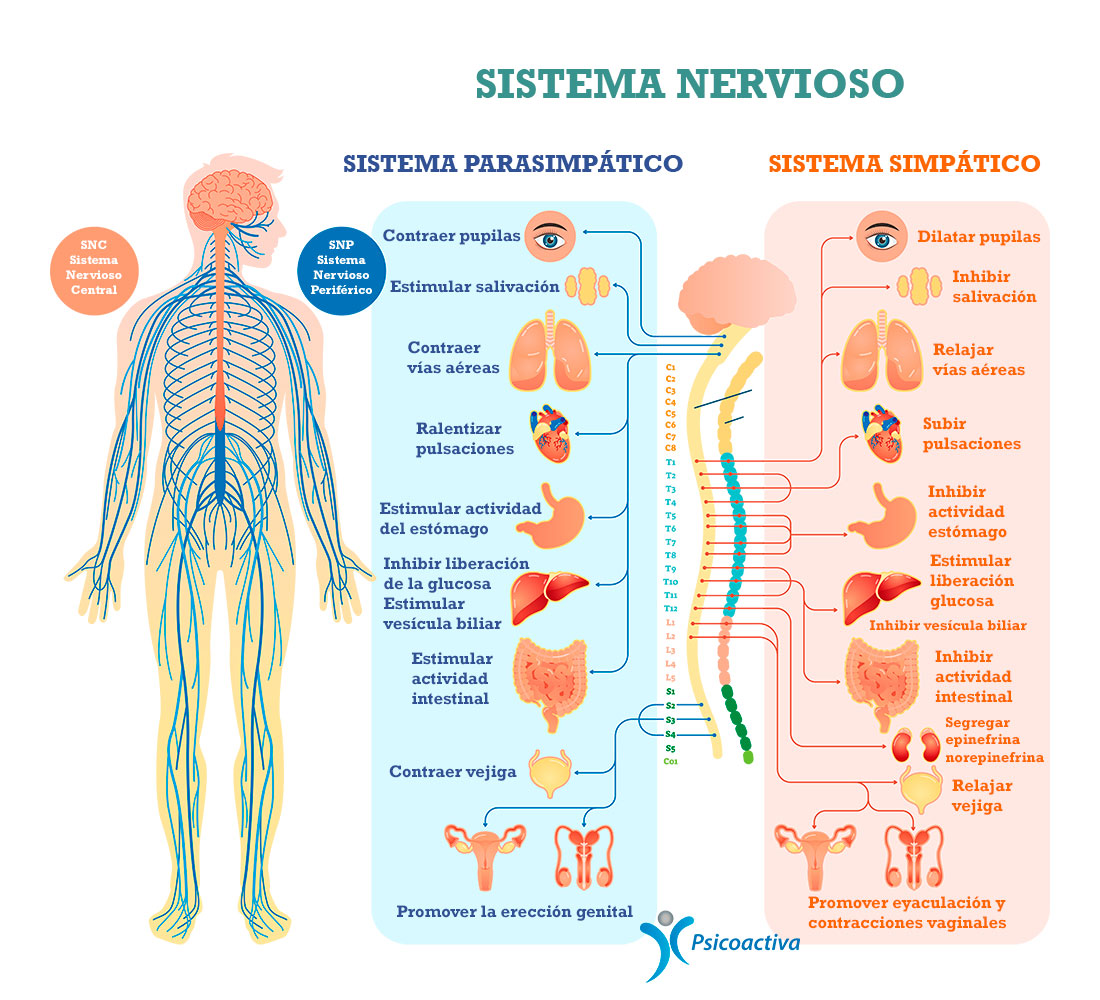
La estimulación simpática causa vasoconstricción y aumenta la frecuencia y aumenta la frecuencia y la actividad de bombeo por el corazón.
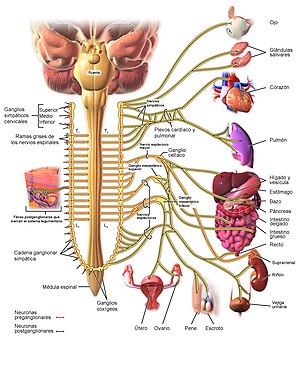
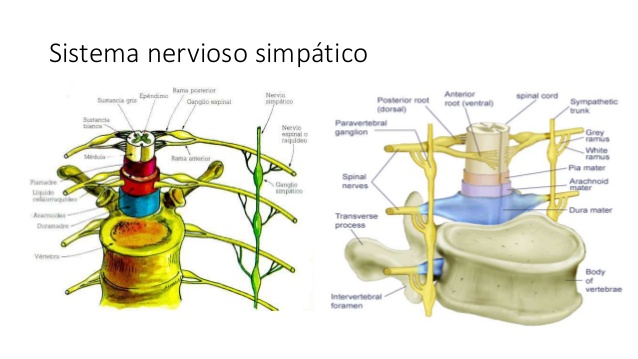
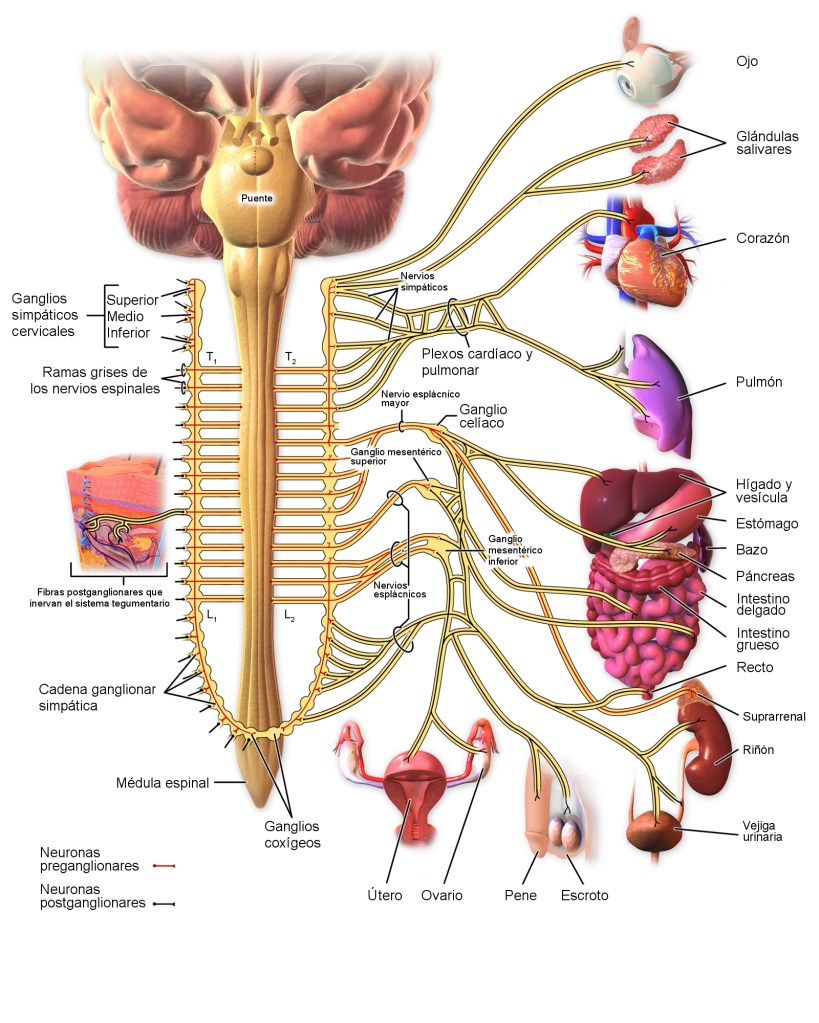
Los nervios salen de la médula espinal a través de los nervios de la columna torácica y algunas lumbares, se dirigen a las cadenas simpáticas para luego seguir dos rutas a la circulación:
A través de los nervios simpáticos que inervan la vasculatura de las vísceras y el corazón, y
En las porciones periféricas de los nervios raquídeos que se distribuyen a la vasculatura de las zonas periféricas. La mayoría de los vasos sanguíneos están inervados por fibras nerviosas simpáticas, a excepción de los capilares. La estimulación simpática aumenta la resistencia vascular y disminuye la velocidad del flujo sanguíneo.
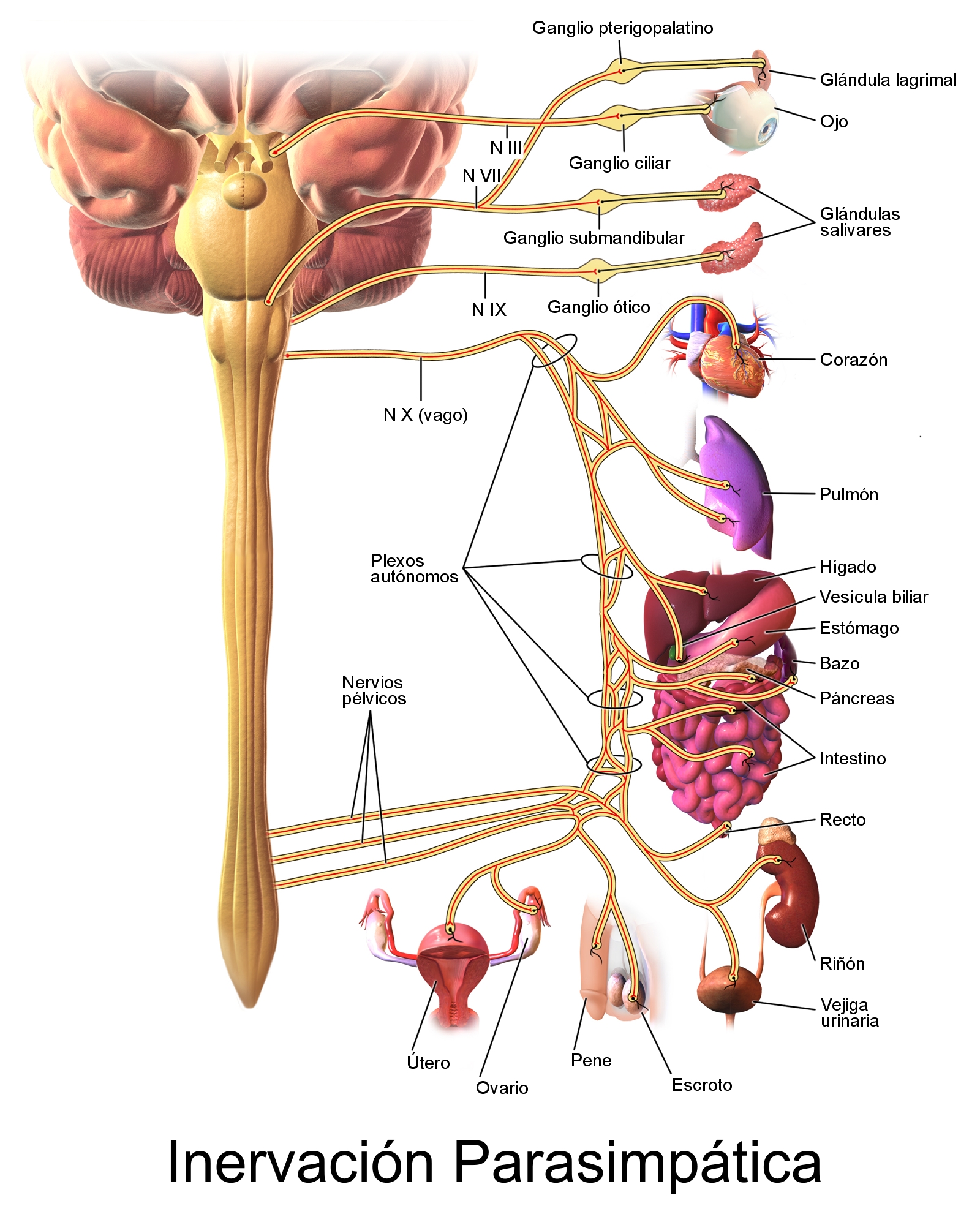
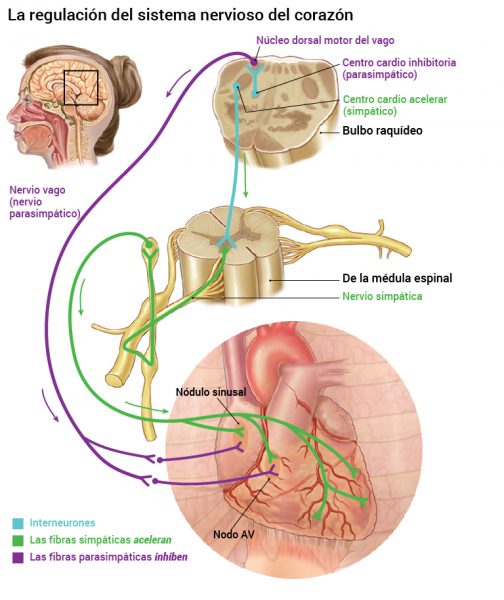
La estimulación parasimpática disminuye la frecuencia y la actividad de bombeo por el corazón.
Su principal función en el control de la circulación consiste en provocar un importante descenso de la frecuencia cardíaca y un pequeño descenso de la contractilidad del músculo cardíaco.
Control del sistema vasoconstrictor simpático por el sistema nervioso central.
Los nervios simpaticos trasnportan una enorme cantidad de fibras nerviosas vasoconstrictoras y solo algunas vasodilatadoras.
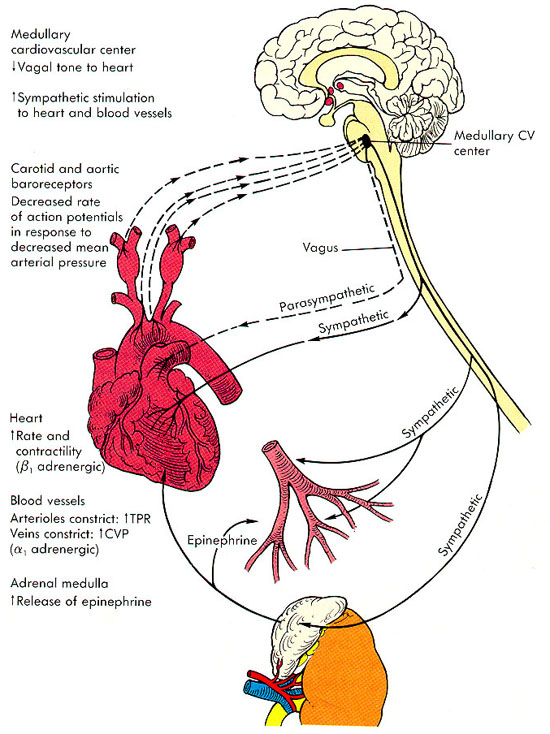
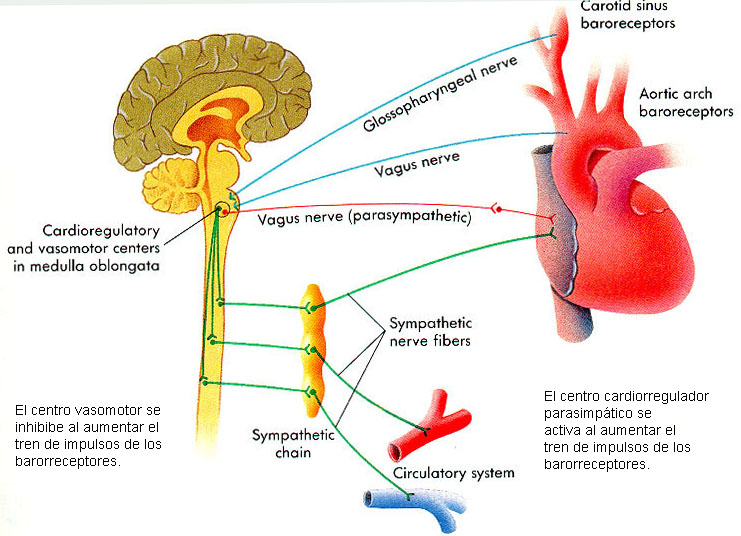
Centro vasomotor del cerebro y control del sistema vasoconstrictor.
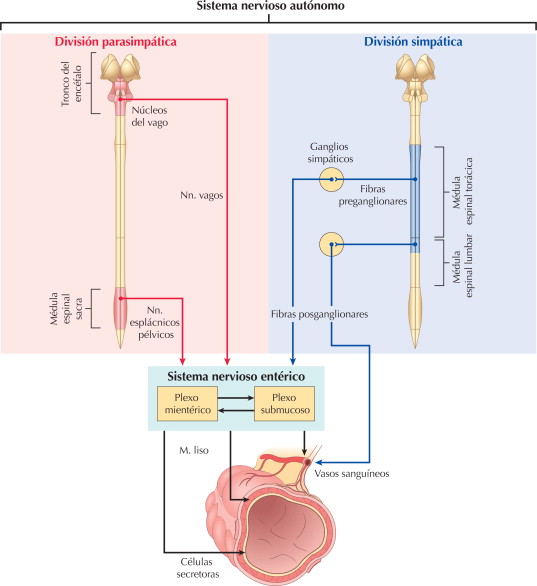
Situado bilateralmente en la sustancia reticular del bulbo y en el tercio inferior de la protuberancia, conforma una zona denominada centro vasomotor. Este centro transmite los impulsos parasimpáticos a través de los nervios vagos hacia el corazón y transmite los impulsos simpáticos a través de la médula espinal y los nervios simpáticos periféricos prácticamente hacia todas las arterias, arteriolas y venas del organismo.
Una zona vasoconstrictora situada bilateralmente en las porciones anterolaterales de la parte superior del bulbo.
Una zona vasodilatadora situada bilateralmente en las porciones anterolaterales de la mitad inferior del bulbo.
Una zona sensitiva situada bilateralmente en los tractos solitarios de las porciones posterolaterales del bulbo y parte inferior de la protuberancia.
Las neuronas de esa zona reciben señales nerviosas sensitivas desde el sistema circulatorio, principalmente a través de los nervios vagos y glosofaríngeo.
El tono simpático vasoconstrictor continuo provoca la construcción de la mayoría de los vasos sanguíneos.
Este tono vasoconstrictor simpático mantiene un estado parcial de la contracción de los vasos sanguíneos. Cuando esta bloqueado por la anestesia raquídea los vasos se dilatan y el presión desciende hasta lo 50 mmHg.
Control del centro vasomotor por los centros nerviosos superiores.
Áreas situadas en el mesencéfalo, sustancia reticular de la protuberancia y el diencéfalo, inhiben o excitan el centro vasomotor. El hipotálamo ejerce potentes reacciones en el centro vasomotor, tanto inhibidoras como excitadras. Partes de la corteza cerebral inhiben o excitan el centro vasomotor.
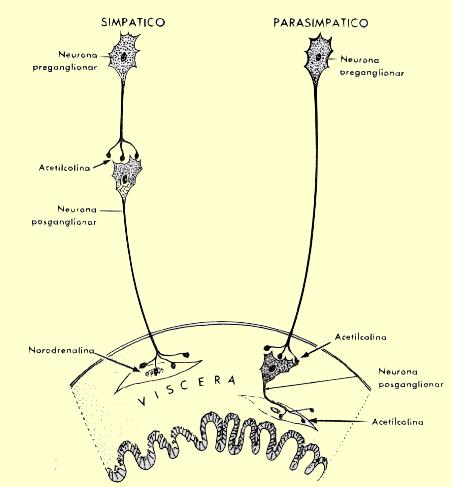
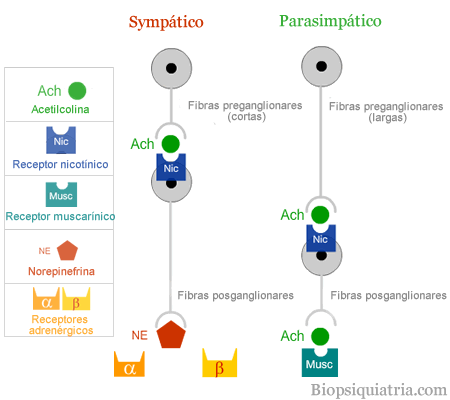
La noradrenalina es el neurotrnasmisor del sistema vasonconstrictor simpático.
Las sustancias de los nervios vasoconstrictores actuan directamente en los receptores a-adrenergéticos del musculo liso vascular, provocando vasoconbstricción.
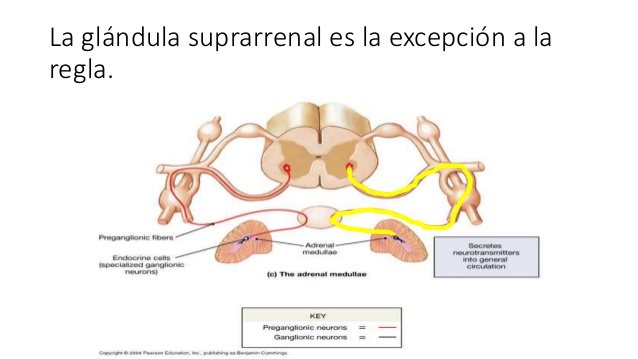
La médula suprarrenal libera noradrenalina y adrenalina durante la estimulación simpática.
Ambas se transportan enbel torrente sanguíneo hacia todas las partes del organismo actuando sobre los vasos , provocando normalmente vasoconstricción al estimular receptores a-adrenérgicos, aunque la adrenalina posee efectos B-adrenérgicos, que provocan vasodilatación en algunos tejidos, como el músculo esquelético.
Función del sistema nervioso en el control rápido de la presión arterial.
Una de las funciones más importante es su capacidad para provocar incrementos rápidos de presión arterial, provocando vasoconstricción y estimulación del corazón. Se producen tres cambios importante simultáneamente, que aumentan la presión al estimular el sistema nervioso autónomo:
Las venas y los demás vasos grandes de la circulación se contraen, lo que desplaza la sangre de los vasos periféricos al corazón y aumenta la fuerza de bombeo, lo que aumenta la fuerza de presión arterial.
La mayoría de lar arteriolas de la circulación sistémica se contraen, la que aumenta la resistencias vascular perfiérica total – presión arterial.
El sistema nervioso autónomo estimula directamente el corazón, lo que potencia la bomba cardíaca.
El sistema nervioso autónomo contribuye al aumento de la presión arterial durante el ejercicio intenso. Se sabe que aumenta de 30-40%. Al mismo tiempo que se activan las zonas motoras cerebrales para iniciar el ejercicio, se activa también la mayor parte del sistema activador reticular del tronco encefálico , incluye una mayor estimulación de zonas vasoconstrictoras y cardioaceleradoras del centro vasommotor. Es instanténeo el aumento de la presión para mantener la sincronización con el ejercicio intenso.
El sistema nervioso autónomo aumenta la presión arterial durante la reacción de alarma.
Cuando se experimenta miedo intenso, la presión arterial aumenta hasta 200 mmHg en seg. Esta “reacción de alarma” proporciona aumento de la presión arterial que puede aportar sangre a cualquier músculo que necesite una respuesta instantánea para huir del peligro.
Mecanismos de reflejo para mantener la presión arterial normal.
El sistema nervioso autónomo actúa para mantener la presión arterial en valores prácticamente normales a través de mecanismos reflejos de retroalimentación negativa.
Sistema de control de la presión arterial mediante barorreceptores.
Básicamente, este reflejo se inicia en los receptores de estiramiento, conocidos como barorreceptores o presorreceptores, situados en puntos específicos de las paredes de varias arterias sistémicas de gran tamaño. Los barorreceptores son muy abundantes en:
Ls pared de ambas arterias carótidas internas, a corta distancia por encima de la bifurcación carotídea (una zona que se conoce como seno carotídeo), y
En la pared del cayado aórtico. Las señales de los «barorreceptores carotídeos» se transmiten a través de los pequeños nervios de Hering, hacia los nervios glosofaríngeos de la parte alta del cuello y después hacia el tracto solitario de la zona del bulbo en el tronco del encéfalo. Las señales que proceden de los «barorreceptores aórticos» del cayado aórtico se transmiten a través de los nervios vagos también hacia el tracto solitario del bulbo.
Los barorreceptores sinusales carotídeos no se estimulan en absoluto con presiones entre 50-60 mmHg, pero en valores superiores responden con una frecuencia progresivamente mayor y alcanzan el máximo en torno a los 180 mmHg. Los barorreceptores responden mucho más a una presión que cambia con gran rapidez que a una presión estacionaria.
Reflejo circulatorio iniciado por los barorreceptores.
Después de que las señales de los barorreceptores entren en el tracto solitario del bulbo, las señales secundarias inhiben el centro vasoconstrictor del bulbo y excitan el centro parasimpático vagal. Los efectos netos son dos;
1) la vasodilatación de las venas y arteriolas en todo el sistema circulatorio periférico y 2) el descenso de la frecuencia cardíaca y de la fuerza de contracción cardíaca.
Función de los barorreceptores durante los cambios de postura del cuerpo.
La capacidad de los barorreceptores de mantener una presión arterial relativamente constante en la parte superior del cuerpo es importante cuando una persona se levanta después de haber estado tumbada.
Función «amortiguadora» de la presión del sistema de control de barorreceptores.
Como el sistema de barorreceptores se opone tanto al aumento como al descenso de la presión arterial, se denomina sistema amortiguador de la presión y los nervios de los barorreceptores se conocen como nervios amortiguadores. Uno de los objetivos principales del sistema arterial de barorreceptores consiste en reducir minuto a minuto la variación de la presión arterial hasta un tercio de la que aparecería si no estuviera presente este sistema.
¿Son importantes los barorreceptores en la regulación a largo plazo de la presión arterial?
Aunque los barorreceptores arteriales proporcionan un control potente de la presión arterial minuto a minuto, su importancia en la regulación a largo plazo de la presión sanguínea es controvertida como consecuencia, tal vez, de que algunos fisiólogos consideran que los barorreceptores tienen una importancia relativamente escasa en la regulación crónica de la presión arterial, porque tienden a reajustarse en 1-2 días a la presión a la cual se exponen.
Control local del flujo sanguíneo en respuesta a las necesidades tisulares
Uno de los principios fundamentales de la función circulatoria es la capacidad de cada tejido de controlar su propio flujo sanguíneo local en proporción a sus necesidades metabólicas. Como son: 1.- Aporte de oxigeno a los tejidos. 2.- Aporte de otros nutrientes, como glucosa, aminoácidos y ácidos grasos. 3.-Eliminaciones de dióxido de carbono de los tejidos. 4.- Eliminaciones de iones hidrogeno de los tejidos. 5.- Mantenimientos de las concentraciones adecuadas de otros iones en los tejidos. 6.- Transporte de varias hormonas y otras sustancias a los distintos tejidos.
Algunos órganos tienen necesidades especiales. Por ejemplo, el flujo sanguíneo de la piel determina la perdida de calor corporal y, de esta forma, se controla la temperatura. Además, el aporte de cantidades adecuadas de plasma sanguíneo a los riñones permite que estos excreten los productos de desecho del organismo
Variaciones de flujo sanguíneo en distintos tejidos y órganos:
Varios cientos de de mililitros por minuto por 100 gramos de tejido tiroideo o suprarrenal y un flujo sanguíneo total de 1350ml/min en el hígado que es de 95 ml/min/100g de tejido hepático.
El flujo sanguíneo tan importante que atraviesa los riñones, 1100ml/min. Esta cantidad tan importante de flujo es necesaria para que los riñones, realicen su función de limpieza de los productos de desecho en la sangre.
en los tejidos en los que la necesidad más importante es la administración de oxigeno, el flujo sanguíneo siempre está controlado a un nivelque solo es ligeramente mayor de lo necesario para mantener la oxigenación tisular plena, pero nada más.
Al controlar el flujo sanguíneo local de una forma tan exacta, los tejidos casi nuncapadecen una deficiencia nutricional de oxigeno y, a pesar de ello.
La carga de trabajo del corazón se mantiene al mínimo.
Mecanismo de control del flujo sanguíneo.
El control de flujo sanguíneo local se puede dividir en las dos fases 1 control a corto plazo y 2, control a largo plazo.
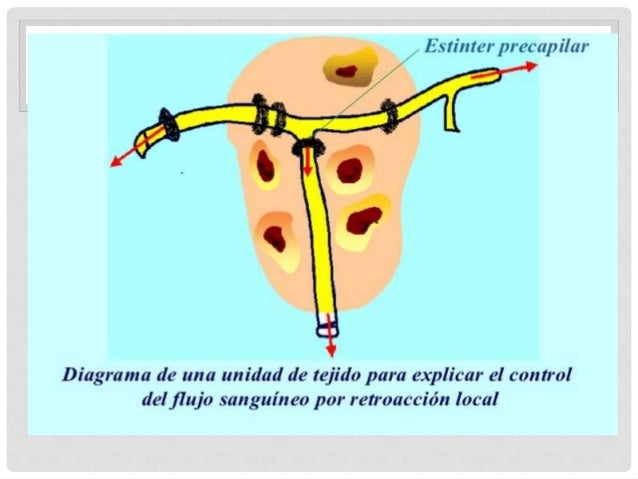
El control a corto plazo se consigue con cambios rápidos de la vasodilatación o vasoconstricción local de las arteriolas, metarteriolas y esfínteres pre capilar, que se producen en segundos o minutos para proporcionar con gran rapidez el mantenimiento del flujo sanguíneo tisular local apropiado.
Hay dos teorías básicas para la regulación del flujo sanguíneo local cuando cambia el metabolismo tisular o disponibilidad de oxigeno:
1.- la teoría vasodilatadora: de la regulación a corto plazo del flujo sanguíneo local: posible papel especial de la adenosina. Según esta teoría, cuanto mayor sea el metabolismo o menor sea la disponibilidad de oxigeno o de algunos otros nutrientes en un tejido, mayor será la velocidad de formación de sustancias vasodilatadoras en las células de ese tejido. Se cree que estas sustancias vasodilatadoras difunden a través de los hacia los esfínteres precapilares, las metarteriolas y las arteriolas para provocar la dilatación. Se han propuesto varias sustancias vasodilatadoras diferentes, como adenosina, dióxido de carbono, compuesto con fosfato de adenosina, histamina, iones potasio e iones hidrogeno.
2.- la teoría de la falta de oxigeno:más exactamente, la teoría de la falta de nutrientes (porque están implicados otros nutrientes, además del oxigeno). El oxigeno (y también otros nutrientes) es necesario como uno de los nutrientes metabólicos para provocar la concentración muscular. Por lo tanto es razonable creer que los vasos sanguíneos simplemente se relajarían en ausencia de una cantidad adecuada de oxigeno, dilatándose de forma natural. Además el aumento del metabolismo podría en teoría disminuir la disponibilidad de oxigeno hacia las fibras musculares lisas de los vasos sanguíneos locales, lo cual también provocaría la vasodilatación local
La mayoría de las teorías vasodilatadoras supone que la sustancia vasodilatadoras se libera del tejido principal menta en respuestas a la deficiencia de oxígeno.
Aproximadamente proporcional a las necesidades de nutrición del tejido .los esfínteres precapilares y las metarteriolas se abren y se cierran cíclicamente varias veces por minuto, siendo proporcional la duración de las fases abiertas a las necesidades metabólicas de oxigeno en los tejidos la apertura y el cierre cíclicos se denominan vasomotilidad.
según los datos disponibles la teoría de la sustancia vasodilatadora o la teoría de la falta de oxigeno explicaría la regulación sanguínea local a corto plazo en respuestas a las necesidades metabólicas de los tejidos. Probablemente, en realidad se produzca una combinación de los dos mecanismos.Posible función de otro nutrientes además del oxigeno en el control del flujo sanguíneo local. En situaciones especiales se ha demostrado que la ausencia de glucosa en la sangre perfundida provoca la vasodilatación tisular local.
Además se produce vasodilatación en una deficiencia de vitaminas que se conoce como beriberi, en la cual el paciente tiene una deficiencia de las sustancias del grupo b tiamina, niacina y riboflavina.
En esta enfermedad el flujo sanguíneo vascular periférico de cualquier parte del cuerpo entre dos y tres veces.Ejemplos especiales de del control “metabólico” a corto plazo del flujo sanguíneo localLos mecanismos que hemos descrito hasta aquí para controlar el flujo sanguíneo local se denominan “mecanismos metabólicos” porque todos ello funcionan en repuestas a las necesidades metabólicas de los tejidos.
Hay otro otros dos ejemplos especiales de control metabólico del flujo sanguíneo local, la hiperemia reactiva y la hiperemia activa.Hiperemia reactiva.- la hiperemia reactiva es otra manifestación del mecanismo deregulación “metabólico” del flujo sanguíneo, es decir, la falta de flujo pone en marcha todos estos factores que provocan la vasodilatación
Regulación a largo plazo del flujo sanguíneo
la mayoría de los mecanismos de regulación del flujo sanguíneo local que hemos comentado actúan en pocos segundos o minutos después del cambio de la situación tisular local.
A pesar de ello, el flujo sanguíneo se ajusta solo en las tres cuartas partes de las necesidades adicionales de los tejidos, incluso después de la activación completa de esos mecanismos agudos.
La regulación a largo plazo del flujo sanguíneo es especialmente importante cuando cambian las demandas metabólicas del tejido a largo plazo.
Es decir, si untejido esta crónicamente hiperactivo y, por tanto requiere u aumento crónico de lascantidades de oxigeno y otros nutrientes, por lo que en alguna semanas aumentantanto el numero como el tamaño de las arteriolas y los vasos capilares para cubrir las necesidades del tejido, a menos que el aparato circulatorio se vuelva patológico o sea demasiado viejo para responder.
las arterias y arteriolas muy pequeñas de cada tejido mediante la deficiencia de sustancias vasodilatadoras o de oxigeno en las células tisulares solo pueden llegara esos vasos, no a las arterias intermedias y mayores del territorio proximal.
Aun así, cuando aumenta el flujo sanguíneo a través de una porción microvascular de la circulación, secundariamente se activa otro mecanismo que dilata también las arterias mayores sintetizan varias sustancias que liberadas afectan el grado de relajación o contracción de la pared arterial.
El flujo de sangre rápido a través de las arterias y arteriolas provoca fuerzas de cizallamiento sobre las células endoteliales.
Regulación a largo plazo del flujo sanguíneo.Hasta este momento, la mayoría de los mecanismos de regulación del flujo sanguíneo local que hemos comentado actúan en pocos segundos o minutos después del cambio de la situación tisular local.
A pesar de ello, el flujo sanguíneo se ajusta solo en las tres cuartas partes de las necesidades adicionales de los tejidos, incluso después de la activación completa de esos mecanismos agudos.
La regulación a largo plazo del flujo sanguíneo es especialmente importante cuando cambian las demandas metabólicas del tejido a largo plazo. Es decir, si untejido esta crónicamente hiperactivo y, por tanto requiere u aumento crónico de lascantidades de oxigeno y otros nutrientes, por lo que en alguna semanas aumentantanto el numero como el tamaño de las arteriolas y los vasos capilares para cubrir las necesidades del tejido, a menos que el aparato circulatorio se vuelva patológico o sea demasiado viejo para responder.
Función del oxigeno en la regulación a largo plazo.
El oxigeno es importante no solo para el control a corto plazo del flujo sanguíneo local, sino también para el control a largo plazo. Un ejemplo es el aumento de la vascularización de los tejidos en los animales que viven en altitudes elevadas, donde el oxigeno atmosférico es bajo. El exceso de oxigeno provoca la interrupción casi inmediata del crecimiento vascular nuevo en la retina de los ojos del niño prematuro incluso provoca la degeneración de algunos de los vaso pequeños que ya se han formado.
Hay una docena o más factores que aumentan el crecimiento de los vasos sanguíneos nuevos, siendo casi todos ellos péptidos pequeños, tres de los mejor identificados son el factor de crecimiento de los fibroblastos, el factor decrecimiento del endotelio vascular (VEGF) y la angiogenina, aislados cada uno de ellos en tejido que tienen un aporte sanguíneo inadecuado. Presumiblemente, es la deficiencia de oxigeno tisular o otros nutrientes la que provoca la formación de los factores de crecimiento vascular (también denominados “factores angiogenicos”)
Cuando se bloquean una arteria o una vena en cualquier tejido del organismo se desarrolla un canal vascular nuevo rodeando el bloqueo y permitiendo que se vuelva a suministrar sangre al tejido afectado, al menos parcialmente. La primera etapa de este proceso es la dilatación de los bucles vasculares pequeños que ya conectan ese vaso `proximal al bloqueo con el vaso distal. Después el flujo es menor de la cuarta parte de lo necesario para cubrir todas las necesidades tisulares.
Los vasos colaterales continúan creciendo durante muchos meses después, casi siempre formando muchos canales colaterales pequeños en lugar de un único vaso de gran tamaño. En reposo, el flujo sanguíneo vuelve muy cerca de los valores normales, pero los nuevos canales son suficientemente grandes como para aportar el flujo sanguíneo necesario durante la actividad tisular agotadora.
Control humoral de la circulación.
El control humoral de la circulación se refiere al control por las sustancias segregadas o absorbidas en los líquidos del organismo, como hormonas e iones.
Sustancias vasoconstrictoras: Noradrenalina, adrenalina, angiotensina II, Vasopresina, Endotelina
Control vascular por iones y otros factores químicos: Hay muchos iones y otros factores químicos que pueden dilatar o contraer los vasos sanguíneos locales.
La mayoría de ellos tiene una función escasa en la regulación global de la circulación, pero hay algunos efectos específicos, como son: 1. El aumento de la concentración del ion calcio provoca vasoconstricción. 2. El aumento de la concentración del ion potasio provoca vasodilatación. 3. El aumento de la concentración del ion magnesio provoca una vasodilatación potente. 4. El aumento de la concentración del ion hidrogeno (descenso de pH) provoca la dilatación de las arteriolas. 5. Los iones acetato y citrato, que provocan una vasodilatación pequeña. 6. El aumento dela concentración de dióxido de carbono provoca una vasodilatación moderada y la mayoría delos tejidos, pero una vasodilatación importante en el cerebro
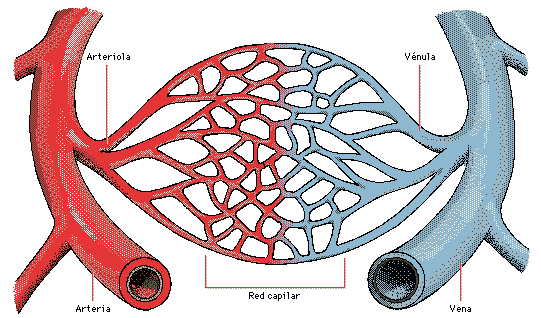
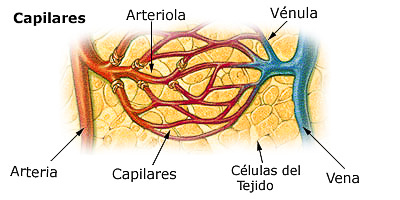
la función circulatoria tiene lugar en los capilares. Los capilares cuentan únicamente con una capa sencilla de células endoteliales altamente permeables, lo que permite un rápido intercambio de nutrientes y de productos celulares de desecho entre los tejidos y la sangre circulante.
La sangre entra en los capilares a través de una arteriola y sale a través de una vénula. La sangre de las arteriolas entra en una serie de metaarteriolas que tienen estructuras entre las arteriolas y los capilares.
Las arteriolas son vasos muy musculares y desempeñan un importante papel en el control del flujo sanguíneo hacia los tejidos.
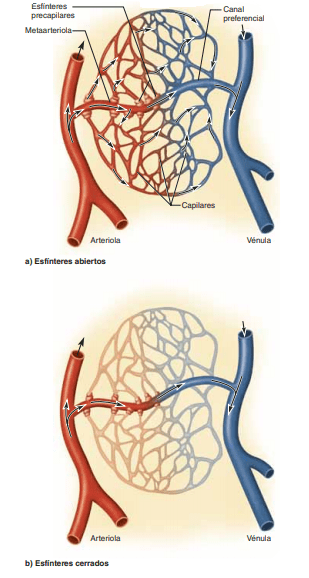
Las metaarteriolas no tienen una capa muscular lisa continua, sino fibras musculares lisas que rodean el vaso en puntos intermitentes, denominados esfínteres precapilares. La contracción del músculo en dichos esfínteres puede abrir y cerrar la entrada al capilar.
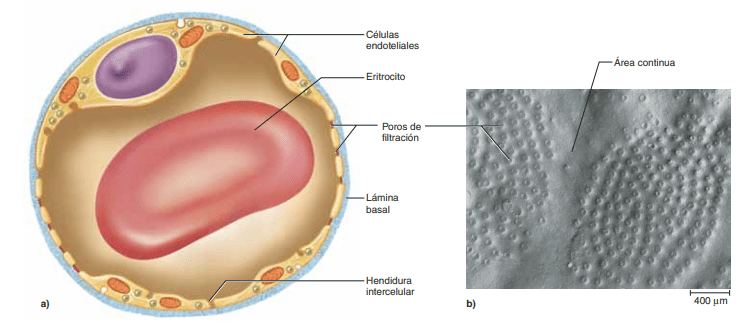
La fina pared capilar consiste en una única capa de células endoteliales. Los capilares son también muy porosos, con varios millones de hendiduras, o poros, entre las células que forman sus paredes en cada centímetro cuadrado de superficie capilar. Dada la elevada permeabilidad de los capilares para la mayoría de los solutos y dada su gran superficie, a medida que el flujo sanguíneo los atraviesa, una gran cantidad de sustancias disueltas se difunde en ambas direcciones a través de esos poros.
Los flujos sanguíneos atraviesan los capilares de forma intermitente, un fenómeno denominado «vasomovilidad». En muchos tejidos, el flujo sanguíneo que atraviesa los capilares no es continuo La causa de esta intermitencia es la contracción intermitente de las metaarteriolas y los esfínteres precapilares, que depende principalmente de la concentración de oxígeno y los productos de desecho del metabolismo tisular.Cuando se reducen las concentraciones de oxígeno en un tejido, los períodos intermitentes del flujo sanguíneo capilar se activan más a menudo y duran más, con lo que se permite que la sangre capilar transporte mayores cantidades de oxígeno y otros nutrientes hacia los tejidos.
la difusión es el medio mas importante por el cual se transfieren las sustancias entre el plasma y el liquido intersticial. a medida que el flujo sanguíneo atraviesa los capilares, un numero enorme de moléculas de agua y partículas disueltas entra y sale a través de la pared capilar, permitiendo la mezcla continua entre el liquido intersticial y el plasma. las sustancias liposolubles, como el oxigeno y el dióxido de carbono, difunden directamente a través de las membranas celulares sin tener que atravesar los poros. las sustancias hidrosolubles, como la glucosa y los electrolítos, se difunden solo a través de los poros intercelulares en la membrana capilar. los principales factores que afectan a la velocidad de difusion a traves de las paredes capilares son:
1.- el tamaño del poro en el capilar: en la mayoría de los capilares, el poro mide 6-7 nm. los poros de algunas membranas capilares, un ejemplo son los sinusoides hepáticos. 2.- el tamaño molecular de la sustancia que se difunde: el agua y la mayoría de los electrolitos, como el sodio y el cloruro, tiene un tamaño molecular menor que el tamaño del poro, lo que permite su rápida difusión a través de la pared capilar, por ejemplo las proteínas que tiene un tamaño molecular mayor que el de los poros. 3.- la diferencia de concentración de la sustancia entre los dos lados de la membrana: cuanto mayor sea la diferencia entre las concentraciones de una sustancia a ambos lados de la membrana capilar, mayor sera la velocidad de difusión en una dirección a través de la membrana.
una sexta parte del volumen total del organismo consiste en espacios entre las células, que colectivamente se conoce como el intersticio. el liquido de estos espacios es el liquido intersticial. el intersticio contiene dos tipos principales de estructuras solidas: a) haces de fibras de colágeno, y b) filamentos de proteoglucano.
El líquido del intersticio deriva por filtración y difusión de los capilares. Contiene casi los mismos componentes que el plasma, excepto por concentraciones mucho más bajas de proteínas, porque las proteínas no atraviesan los poros de los capilares. El líquido intersticial queda atrapado principalmente en los diminutos espacios que hay entre los filamentos de proteoglucanos. Esta combinación de filamentos de proteoglucano y líquido atrapado dentro de ellos tiene las características de un gel y, por tanto, se conoce como gel tisular.
Aunque casi todo el líquido del intersticio está atrapado dentro del gel tisular, a veces también hay pequeños riachuelos de líquido «libre» y pequeñas vesículas de líquido libre, lo que significa que carece de moléculas de proteoglucano y, por tanto, puede fluir libremente.
El sistema linfático también tiene su importancia, al devolver a la circulación las pequeñas cantidades del exceso de proteína y líquido que se pierde desde la sangre hacia los espacios intersticiales.
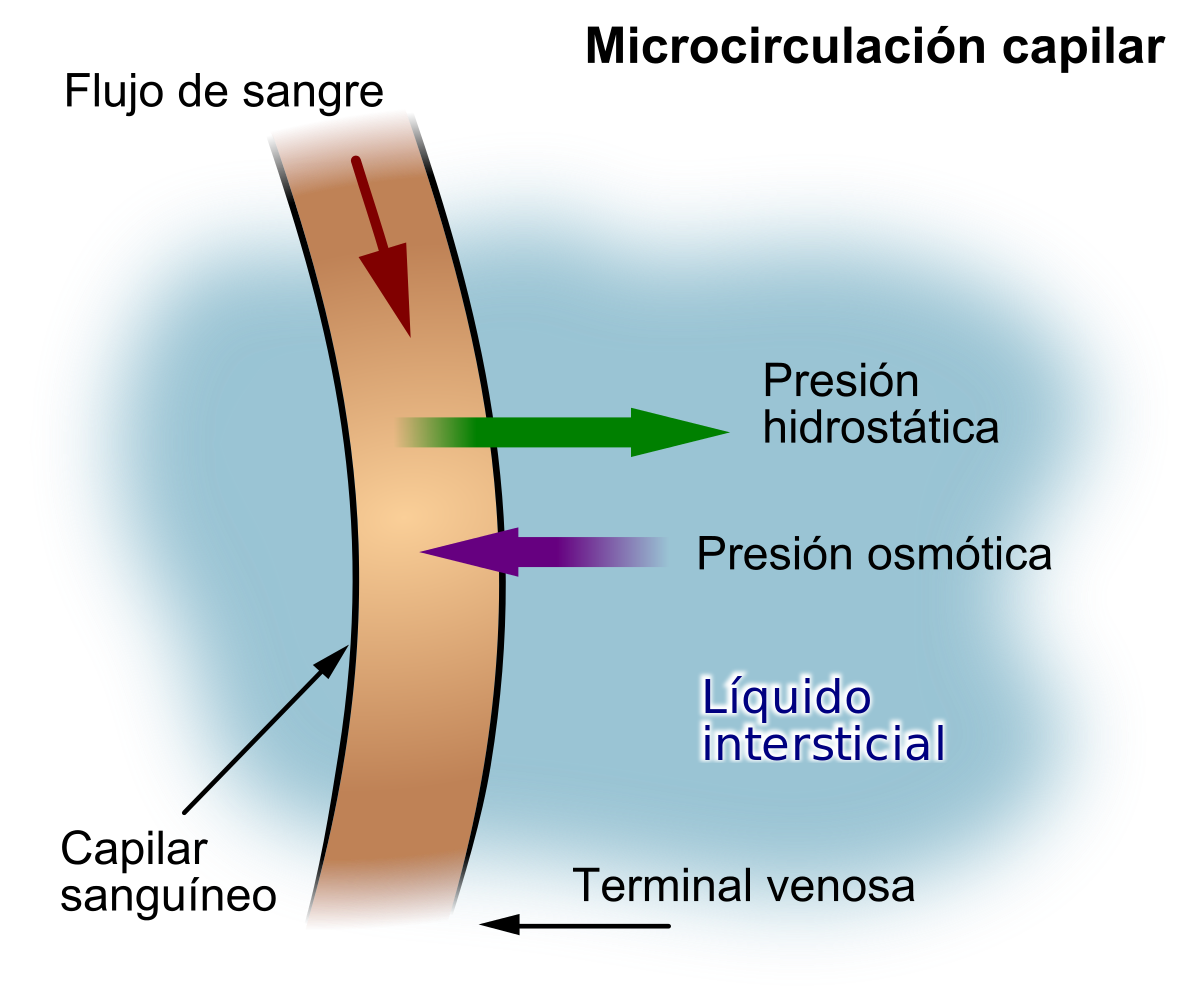
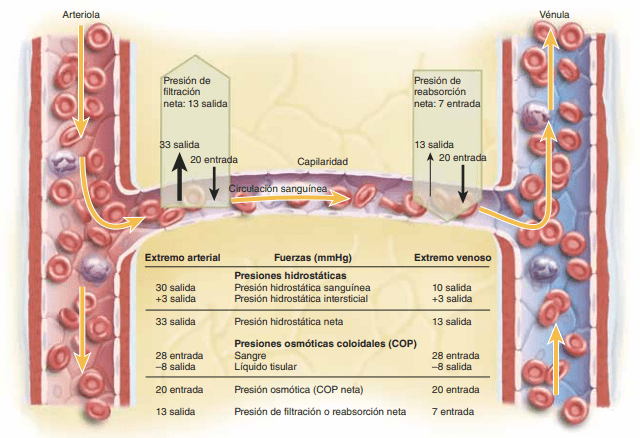
«fuerzas de Starling» en honor al fisiólogo Ernest Starling, que demostró su importancia por primera vez, son:
La presión capilar (Pc), que tiende a forzar la salida del líquido a través de la membrana capilar.
La presión del líquido intersticial (Pif), que tiende a forzar la entrada del líquido a través de la membrana capilar cuando la Pif es positiva, pero fuerza la salida cuando la Pif es negativa.
La presión coloidosmótica del plasma en el capilar (Πp), que tiende a provocar ósmosis de líquido hacia el interior a través de la membrana capilar.
La presión coloidosmótica del líquido intersticial (Πif), que tiende a provocar la ósmosis del líquido hacia el exterior a través de la membrana capilar.
El sistema linfático representa una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. Es más, los vasos linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de estas podrá ser eliminada por absorción directamente hacia los capilares sanguíneos. Este retorno de las proteínas a la sangre desde los espacios intersticiales es una función esencial sin la cual moriríamos en 24 h.
La mayoría del líquido que se filtra desde los extremos arteriales de los capilares sanguíneos fluye entre las células y, por último, se reabsorbe de nuevo hacia los extremos venosos de los capilares sanguíneos; pero, como media, aproximadamente la décima parte del líquido entra en los capilares linfáticos y vuelve hacia la sangre a través del sistema linfático y no al contrario, a través de los capilares venosos. La cantidad total de toda esta linfa normalmente es de solo 2-3 l al día.
La linfa deriva del líquido intersticial que fluye en los linfáticos, por lo que la linfa que entra primero en los vasos linfáticos terminales tiene casi la misma composición que el líquido intersticial. La concentración de proteínas en el líquido intersticial de la mayoría de los tejidos alcanza un promedio de 2 g/dl y la concentración de proteínas del flujo linfático que procede de estos tejidos es aproximada a este valor. La linfa formada en el hígado tiene una concentración de proteínas hasta de 6 g/dl y la linfa formada en el intestino tiene una concentración de proteínas hasta de 3-4 g/dl. Como aproximadamente dos tercios de toda la linfa procede normalmente del hígado y los intestinos, la linfa del conducto torácico, que es una mezcla de linfa de todas las áreas del organismo, tiene una concentración de proteínas en torno a 3-5 g/dl. El sistema linfático también es una de las vías principales de absorción de los nutrientes del aparato digestivo.
Todos los vasos sanguíneos del aparato vascular son distensibles, esto permite el acoplamiento de las arterias
al gasto pulsátil del corazón y superar las pulsaciones de la presión. Los vasos más distensibles del cuerpo son las venas, capaces de almacenar 0.5-1 litro de sangre. Las arterias son más fuertes que las venas, es por eso que estas últimas son más distensibles. La estimulación simpática aumenta la presión en cada volumen de arterias o venas, mientras que la inhibición simpática lo disminuye. Compliancia diferida: se refiere a que si un vaso está expuesto a un aumento de volumen, primero presentara un incremento de la presión pero progresivamente se estirara el musculo liso del vaso y hará que la presión vuelva a la normalidad.
Pulsaciones de la presión arterial
La compliancia del árbol arterial permite reducir las pulsaciones de la presión hasta que prácticamente desaparecen al momento de alcanzar los capilares.
Presión sistólica -> 120 mmHg ; presión diastólica -> 80 mmHg la diferencia entre estas dos presiones recibe el nombre de presión de pulso y es igual a -> 40mmHg.
Factores que afectan la presión de pulso:
Volumen sistólico del corazón.
Compliancia (distensibilidad total) del árbol arterial (en los ancianos la distensibilidad de los vasos disminuye por la arterioesclerosis).
Eyección del corazón durante la sístole.
La presión de pulso está determinada por la relación entre el gasto cardíaco y la compliancia del árbol arterial.
Situaciones que provocan un perfil anormal de la onda de pulso de presión:
Estenosis valvular aórtica: se debe a la disminución del diámetro de apertura de la válvula lo cual la presión de pulso aórtico disminuye también.
Conducto arterioso permeable: es cuando la sangre en lugar de bombearse a la aorta fluye a través de la arteria y vasos pulmonares, lo que produce un descenso diastólica antes del latido.
Insuficiencia aórtica: es por la ausencia o por el cierre incompleto de la válvula, lo que produce que después de cada latido la sangre que fluye por la aorta se va hacia el ventrículo izquierdo, lo que provoca produce una disminución de la presión aortica.
La velocidad de la transmisión del pulso de la presión en la aorta normal es de 3-5 m/s, de 7-10 m/s en las ramas arteriales grandes y de 15-35 m/s en las pequeñas arterias.
La disminución de las pulsaciones en la periferia recibe el nombre de amortiguación de los pulsos, sus orígenes son:
Resistencia al movimiento de la sangre en los vasos.
Compliancia de los vasos.
El grado de amortiguación es casi directamente proporcional al producto resistencia por compliancia.
Se utiliza la arteria antecubital en el método de auscultación para determinar las presiones arteriales. Los ruidos que se escucha reciben el nombre de ruidos de Korotkoff.
Las venas y sus funciones
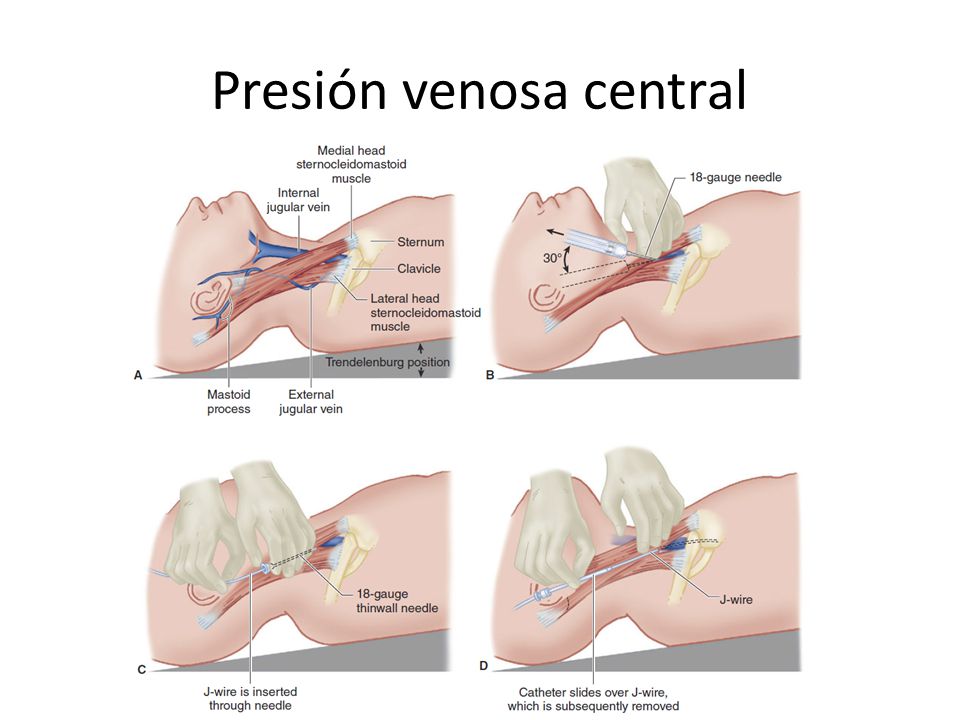
Las venas periféricas pueden impulsar la sangre mediante la bomba venosa e incluso pueden regular el gasto cardíaco. La sangre de todas las venas sistémicas fluyen hacia la aurícula izquierda del corazón. La presión del interior de esta cámara recibe el nombre de presión venosa central.
La presión de la aurícula izquierda está regulada por:
La capacidad del corazón de bombear la sangre al exterior de la aurícula y el ventrículo derecho a los pulmones.
La tendencia de la sangre a fluir desde las venas periféricas hacia la aurícula derecha.
Factores que aumentan el retorno venoso:
Aumento del volumen de sangre.
Aumento del tono de los grandes vasos del organismo.
Dilatación de las arteriolas.
La presión normal de la aurícula derecha es de 0 mmHg.
Cuando la presión intraabdominal aumenta, la presión de las venas de las piernas debe de aumentar por encima de la presión abdominal antes de que las venas abdominales se abran y permitan el paso de la sangre desde las piernas al corazón.
Presión gravitacional o hidrostática: es producida en el aparato vascular por el peso de la sangre en las venas.
Las venas del cuello de una persona que esté de pie se colapsan casi por completo en todo su recorrido hasta el cráneo, por la presión atmosférica que hay fuera del cuello.
Las venas del interior del cráneo se encuentran dentro de una cámara no colapsable (la cavidad craneal), por lo que no se pueden colapsar. En consecuencia, puede haber una presión negativa en los senos de la dura de la cabeza; en bipedestación la presión venosa del seno sagital de la parte superior del cráneo es de –10 mmHg, por la “aspiración” hidrostática que existe entre la parte superior y la base del cráneo.
Las válvulas de las venas están distribuidas a tal manera que la dirección del flujo sanguíneo venoso solo puede ir hacia el corazón.
Bomba venosa o bomba muscular: es cuando la tensión de los músculos empuja una determinada cantidad de sangre venosa al corazón.
La presión venosa puede determinarse observando el grado de distensión de las venas periféricas, especialmente la del cuello, o de una manera exacta a través de un catéter a través de las venas periféricas hasta la aurícula derecha.
La presión medida de la aurícula derecha es de 0mmHg y la presión arterial es de 100mmHg.
En la válvula tricúspide o cerca de ella no afectan los factores de presión gravitacional. El corazón actúa como un regulador de retroalimentación de presión en la válvula tricúspide. Más del 60% de toda la sangre venosa se encuentra en las venas, es por eso que se dice que actúan como reservorio de energía.
Reservorios sanguíneos específicos:
Bazo -> puede liberar hasta 100 ml de sangre (pulpa roja).
Hígado -> puede liberar unos cientos litros de sangre.
Venas abdominales grandes -> liberan hasta 300 ml de sangre. ->Plexos venosos situados en la piel -> liberan unos cientos de ml de sangre.
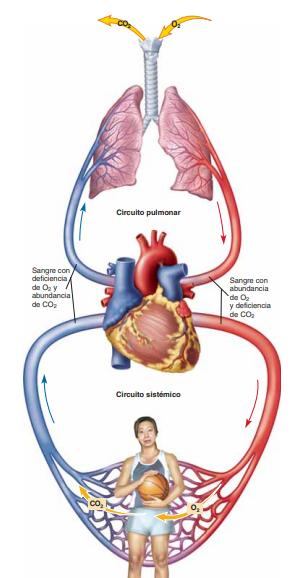
La función de la circulación consiste en atender las necesidades del organismo: transportar nutrientes hacia los tejidos del organismo, transportar los productos de desecho, transportar las hormonas de una parte del organismo a otra y, en general, mantener un entorno apropiado en todos los líquidos tisulares del organismo para lograr la supervivencia y funcionalidad óptima de las células. Está divida en circulación sistémica y circulación pulmonar:
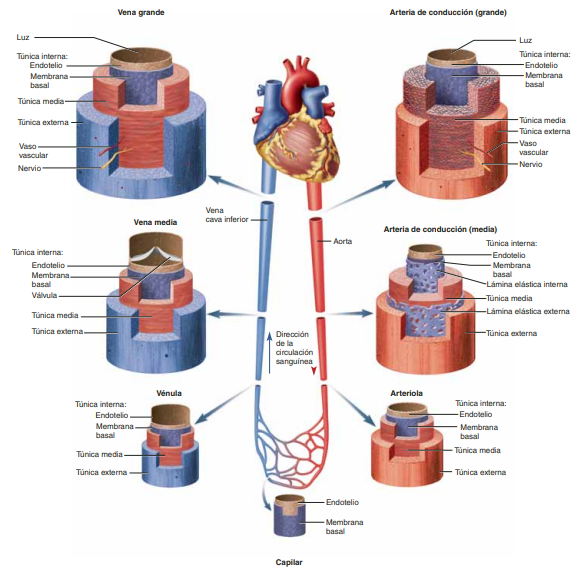
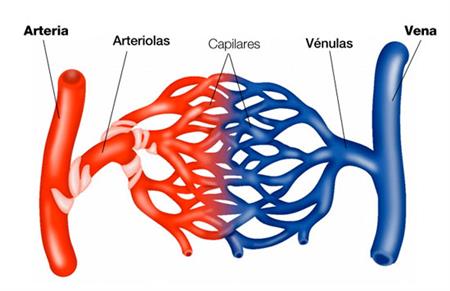
La función de las arterias consiste en transportar la sangre con una presión alta hacia los tejidos, motivo por el cual las arterias tienen unas paredes vasculares fuertes y unos flujos sanguíneos importantes con una velocidad alta. Las arteriolas son las últimas ramas pequeñas del sistema arterial y actúan controlando los conductos a través de los cuales se libera la sangre en los capilares.
Tienen paredes musculares fuertes que pueden cerrarlas por completo o que pueden, al relajarse, dilatarlos vasos varias veces.Los capilares consiste en el intercambio de líquidos, nutrientes, electrólitos, hormonas y otras sustancias en la sangre y en el líquido intersticial. Las paredes del capilar son muy finas y tienen muchos poros capilares diminutos, que son permeables al agua y a otrasmoléculas pequeñas.
Las vénulas recogen la sangre de los capilares y después se reúnen gradualmente formando venas de tamaño progresivamente mayor.
Las venas funcionan como conductos para el transporte de sangre que vuelve desde las vénulas al corazón; igualmente importante es que sirven como una reserva importante desangre extra. Paredes finas.
Volúmenes de sangre en los distintos componentes de la circulación.
Superficies transversales y velocidades del flujo sanguíneo:
Volúmenes de sangre en los distintos componentes de la circulación. Superficies transversales y velocidades del flujo sanguíneo Como debe pasar el mismo volumen de flujo sanguíneo (F) a través de cada segmento dela circulación en cada minuto, la velocidad del flujo sanguíneo (v) es inversamente proporcional a la superficie transversal vascular (A).
v=F/A
En condiciones de reposo la velocidad es como media de 33cm/s en la aorta pero con una velocidad sólo de 1/1.000 en los capilares, es decir, aproximadamente 0,3 mm/s. No obstante, como los capilares tienen una longitud de sólo 0,3 a l mm, la sangre sólo se queda allí durante 1-3s.
Presiones en las distintas porciones de la circulación.
La presión arterial alterna entre una presión sistólica de 120 mmHg y una diastólica de 80mmHg, La presión de los capilares sistémicos oscila desde 35 mmHg cerca de los extremos arteriolar es hasta tan sólo 10 mmHg cerca de los extremos venosos, pero la presión media «funcional» en la mayoría de los lechos vasculares es de 17 mmHg, aproximadamente, una presión suficientemente baja que permite pequeñas fugas de plasma a través de los poros diminutos de las paredes capilares, aunque los nutrientes pueden difundir fácilmente a través de los mismos poros hacia las células de los tejido sexternos. La presión sistólica arterial pulmonar alcanza un promedio de 25 mmHg y la diastólica, de 8 mmHg, con una presión arterial pulmonar media de sólo 16 mmHg. La media de la presión capilar pulmonar alcanza un promedio de sólo 7 mmHg. Aun así, el flujo sanguíneo por minuto a través de los pulmones es el mismo que en la circulación sistémica.
Principios básicos de la función circulatoria
1. La velocidad del flujo sanguíneo en cada tejido del organismo casi siempre se controla por precisión en relación con las necesidades del tejido: Hormonas y sistema nervioso colaboran en el control del flujo sanguíneo tisular. 2. El gasto cardíaco se controla principalmente por la suma de todos los flujos tisulares locales. 3. La regulación de la presión arterial es generalmente independiente del control del flujo sanguíneo local o del control del gasto cardíaco.
Interrelaciones entre la presión, el flujo y la resistencia.
El flujo sanguíneo que atraviesa un vaso sanguíneo está determinado por dos factores: 1)diferencia de presión de la sangre entre los dos extremos de un vaso, también denominado «gradiente de presión» en el vaso, que es la fuerza que empuja la sangre a través del vaso 2) los impedimentos que el flujo sanguíneo encuentra en el vaso,que se conoce como resistencia vascular.
El flujo atraves del vaso se puede calcular con la ley de Ohm que es: ° F=∆P/R ° F= Flujo sanguíneo ° ∆P= Diferencia de presiones de los dos extremos del vaso ° R= resistencia
Flujo sanguíneo
Es la cantidad de sangres que atraviesa un punto dado de la circulación en un periodo de tiempo determinado. ml/m o ml/s.
El flujo sanguíneo global de toda la circulación de un adulto en reposo es de unos 5.000ml/min se considera igual al gasto cardiaco porque es la cantidad de sangre que bombea el corazón en la aorta en cada minuto.
Presión sanguínea
Se mide casi siempre en milímetros de mercurio (mmHg) porque el manómetro de mercurio se ha usado como patrón de referencia para medir la presión
La presión arterial mide la fuerza ejercida por la sangre contra una unidad de superficie dela pared del vaso.En ocasiones, la presión se mide en centímetros de agua (cm H20).
Una presión de 10cm H20 significa una presión suficiente para elevar una columna de agua contra la gravedad hasta una altura de 10 centímetros.
Una presión de 1 mm de mercurio es igual a una presión de 1,36 cm de agua.
Presiones gaseosas en una mezcla de gases: presiones parciales de gases individuales
La presión está producida por múltiples impactos de partículas en movimiento contra una superficie. Por tanto, la presión de un gas que actúa sobre las superficies de las vías aéreas y de los alvéolos es proporcional a la suma de las fuerzas de los impactos de todas las moléculas de ese gas que chocan contra la superficie en cualquier momento dado. Esto significa que la presión es directamente proporcional a la concentración de las moléculas del gas. se manejan muestras de gases mezclas de gases, principalmente oxígeno, nitrógeno y dióxido de carbono. La velocidad de difusión de cada uno de estos gases es directamente proporcional a la presión que genera ese gas solo, que se denomina presión parcial de ese gas.
Considérese el aire, que tiene una composición aproximada del 79% de nitrógeno y el 21% de oxígeno. La presión total de esta mezcla al nivel del mar es en promedio de 760 mmHg. A partir de la descripción previa de la base molecular de la presión es evidente que cada uno de los gases contribuye a la presión total en proporción directa a su concentración
Presiones de gases disueltos en agua y tejidos Los gases disueltos en agua o en los tejidos corporales también ejercen una presión, porque las moléculas de gas disuelto se mueven de manera aleatoria y tienen energía cinética. Además, cuando el gas disuelto en el líquido entra en contacto con una superficie, como la membrana de una célula, ejerce su propia presión parcial de la misma manera que un gas en la fase gaseosa.
Factores que determinan la presión parcial de un gas disuelto en un líquido La presión parcial de un gas en una solución está determinada no solo por su concentración, sino también por el coeficiente de solubilidad del gas. Es decir, algunos tipos de moléculas, especialmente el CO2 son atraídas física o químicamente por las moléculas de agua, mientras que otros tipos de moléculas son repelidas. Cuando las moléculas son atraídas se pueden disolver muchas más sin generar un exceso de presión parcial en el interior de la solución. Por el contrario, en el caso de moléculas que son repelidas se generará una presión parcial elevada con menos moléculas disueltas
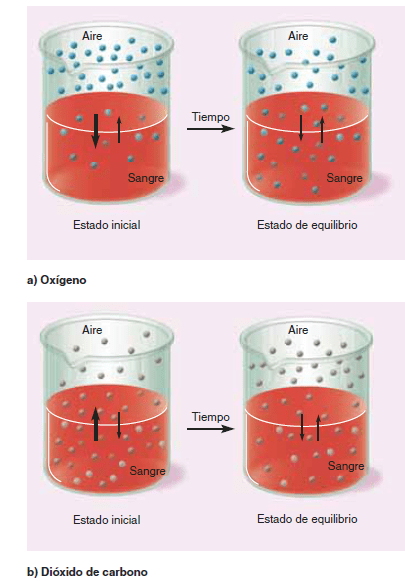
Difusión de gases entre la fase gaseosa de los alvéolos y la fase disuelta de la sangre pulmonar La presión parcial de cada uno de los gases en la mezcla de gas respiratorio alveolar tiende a hacer que las moléculas de ese gas se disuelvan en la sangre de los capilares alveolares. Por el contrario, las moléculas del mismo gas que ya están disueltas en la sangre están rebotando de manera aleatoria en el líquido de la sangre, y algunas de estas moléculas que rebotan escapan de nuevo hacia los alvéolos.
La velocidad a la que escapan es directamente proporcional a su presión parcial en la sangre. la difusión neta está determinada por la diferencia entre las dos presiones parciales. Si la presión parcial es mayor en la fase gaseosa de los alvéolos, como ocurre normalmente en el caso del oxígeno, entonces más moléculas difundirán hacia la sangre que en la otra dirección. Por otro lado, si la presión parcial del gas es mayor en el estado disuelto en la sangre, como ocurre normalmente en el caso del CO2 , la difusión neta se dirigirá hacia la fase gaseosa de los alvéolos.
Presión de vapor de agua Cuando se inhala aire no humidificado hacia las vías aéreas, el agua se evapora inmediatamente desde las superficies de estas vías aéreas y humidifica el aire. Esto se debe al hecho de que las moléculas de agua, al igual que las moléculas de los diferentes gases disueltos, están escapando continuamente de la superficie del agua hacia la fase gaseosa. La presión parcial que ejercen las moléculas de agua para escapar a través de la superficie se denomina la presión de vapor del agua. A la temperatura corporal normal, 37 °C, esta presión de vapor es de 47 mmHg. Por tanto, una vez que la mezcla de gases se ha humidificado totalmente (es decir, una vez que está en «equilibrio» con el agua), la presión parcial del vapor de agua en la mezcla de gases es de 47 mmHg
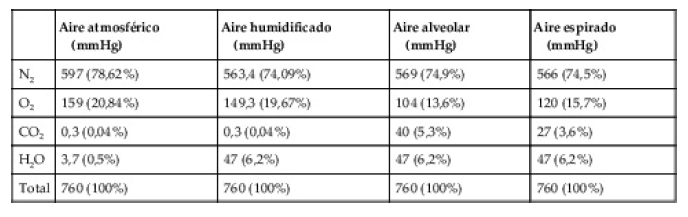
Las composiciones del aire alveolar y el aire atmosférico son diferentes
El aire alveolar no tiene en modo alguno las mismas concentraciones de gases que el aire atmosférico . Hay varias razones para estas diferencias. Primero, el aire alveolar es sustituido solo de manera parcial por aire atmosférico en cada respiración. Segundo, el O2 se absorbe constantemente hacia la sangre pulmonar desde el aire pulmonar. Tercero, el CO2 está difundiendo constantemente desde la sangre pulmonar hacia los alvéolos. Y cuarto, el aire atmosférico seco que entra en las vías aéreas es humidificado incluso antes de que llegue a los alvéolo
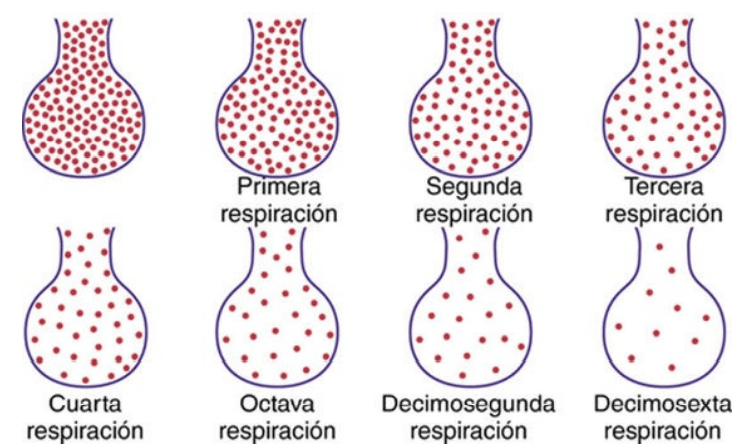
El aire alveolar se renueva lentamente por el aire atmosférico
en promedio la capacidad residual funcional de los pulmones (el volumen de aire que queda en los pulmones al final de una espiración normal) en un hombre mide aproximadamente 2.300 ml. Sin embargo, solo 350 ml de aire nuevo entran en los alvéolos en cada inspiración normal y se espira esta misma cantidad de aire alveolar. Por tanto, el volumen de aire alveolar que es sustituido por aire atmosférico nuevo en cada respiración es de solo 1/7 del total, de modo que son necesarias múltiples inspiraciones para intercambiar la mayor parte del aire alveolar
Concentración y presión parcial de oxígeno en los alvéolos
El oxígeno se absorbe continuamente desde los alvéolos hacia la sangre de los pulmones, y continuamente se respira O2 nuevo hacia los alvéolos desde la atmósfera. Cuanto más rápidamente se absorba el O2, menor será su concentración en los alvéolos; por el contrario, cuanto más rápidamente se inhale nuevo O2 hacia los alvéolos desde la atmósfera, mayor será su concentración. Por tanto, la concentración de O2 en los alvéolos, y también su presión parcial, está controlada por: 1) la velocidad de absorción de O2 hacia la sangre, y 2) la velocidad de entrada de O2 nuevo a los pulmones por el proceso ventilatorio.
El aire espirado es una combinación de aire del espacio muerto y aire alveolar
La composición global del aire espirado está determinada por: 1) la cantidad del aire espirado que es aire del espacio muerto, y 2) la cantidad que es aire alveolar. las modificaciones progresivas de las presiones parciales de O2 y de CO2 en el aire espirado durante el transcurso de la espiración. La primera porción de este aire, el aire del espacio muerto de las vías aéreas respiratorias, es aire humidificado típico, como se muestra en la tabla 40-1. Después cada vez más aire alveolar se mezcla con el aire del espacio muerto hasta que finalmente se ha eliminado el aire del espacio muerto y solo se espira aire alveolar al final de la espiración.
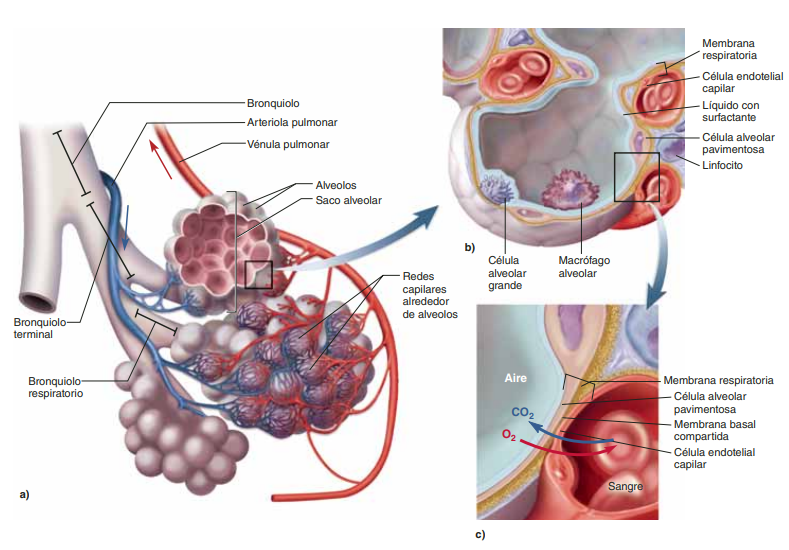
Difusión de gases a través de la membrana respiratoria
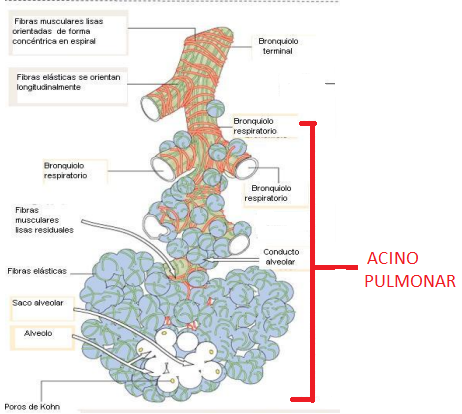
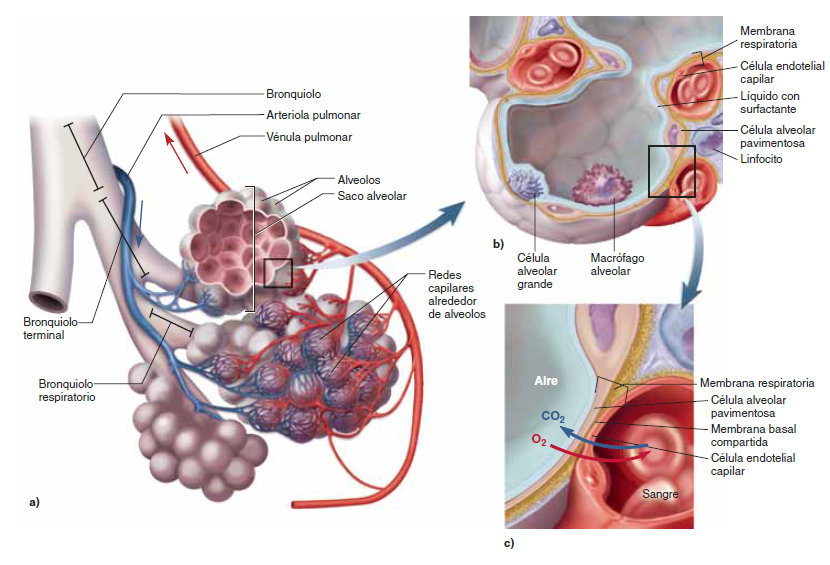
Unidad respiratoria la unidad respiratoria (también denominada «lobulillo respiratorio»), está formada por un bronquíolo respiratorio, los conductos alveolares, los atrios y los alvéolos. Hay aproximadamente 300 millones de alvéolos en los dos pulmones, y cada alvéolo tiene un diámetro medio de aproximadamente 0,2 mm. Las paredes alveolares son muy delgadas y entre los alvéolos hay una red casi sólida de capilares interconectados
Así, es evidente que los gases alveolares están muy próximos a la sangre de los capilares pulmonares. Además, el intercambio gaseoso entre el aire alveolar y la sangre pulmonar se produce a través de las membranas de todas las porciones terminales de los pulmones, no solo en los alvéolos. Todas estas membranas se conocen de manera colectiva como la membrana respiratoria, también denominada membrana pulmonar
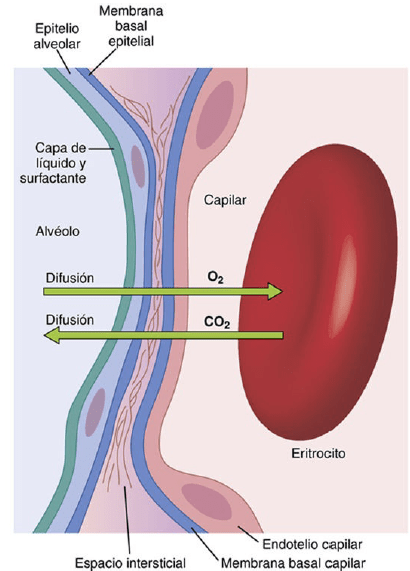
Membrana respiratoria
Se pueden observar las siguientes capas de la membrana respiratoria:
Una capa de líquido que contiene surfactante y que tapiza el alvéolo, lo que reduce la tensión superficial del líquido alveolar.
El epitelio alveolar, que está formado por células epiteliales delgadas.
Una membrana basal epitelial.
Un espacio intersticial delgado entre el epitelio alveolar y la membrana capilar.
Una membrana basal capilar que en muchos casos se fusiona con la membrana basal del epitelio alveolar.
La membrana del endotelio capilar.
A partir de estudios histológicos se ha estimado que el área superficial total de la membrana respiratoria es de aproximadamente 70 m2 en el hombre adulto sano, que es equivalente al área del suelo de una habitación de 7 × 10 m. La cantidad total de sangre en los capilares de los pulmones en cualquier instante dado es de 60 a 140 ml.
Factores que influyen en la velocidad de difusión gaseosa a través de la membrana respiratoria
los factores que determinan la rapidez con la que un gas atraviesa la membrana son: 1) el grosor de la membrana; 2) el área superficial de la membrana; 3) el coeficiente de difusión del gas en la sustancia de la membrana, y 4) la diferencia de presión parcial del gas entre los dos lados de la membrana
Capacidad de difusión de la membrana respiratoria
La capacidad de la membrana respiratoria de intercambiar un gas entre los alvéolos y la sangre pulmonar se expresa en términos cuantitativos por la capacidad de difusión de la membrana respiratoria, que se define como el volumen de un gas que difunde a través de la membrana en cada minuto para una diferencia de presión parcial de 1 mmHg.
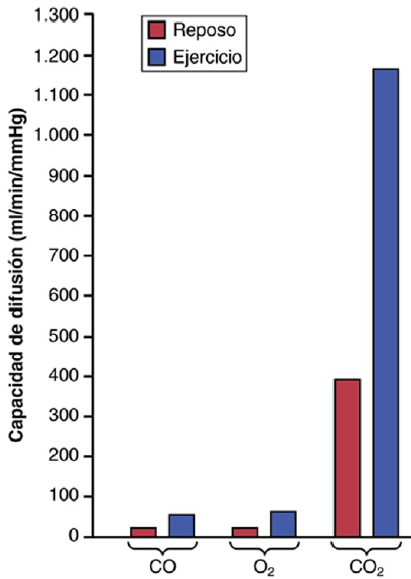
Capacidad de difusión del oxígeno En un hombre joven medio, la capacidad de difusión del O2 en condiciones de reposo es en promedio de 21 ml/min/mmHg. La diferencia media de presión de O2 a través de la membrana respiratoria durante la respiración tranquila normal es de aproximadamente 11 mmHg. La multiplicación de esta presión por la capacidad de difusión (11 × 21) da un total de aproximadamente 230 ml de oxígeno que difunden a través de la membrana respiratoria cada minuto, que es igual a la velocidad a la que el cuerpo en reposo utiliza el O2.
Aumento de la capacidad de difusión del oxígeno durante el ejercicio Durante el ejercicio muy intenso u otras situaciones que aumentan mucho el flujo sanguíneo pulmonar y la ventilación alveolar, la capacidad de difusión del O2 aumenta en los hombres jóvenes hasta un máximo de aproximadamente 65 ml/min/mmHg, que es el triple de la capacidad de difusión en situación de reposo. Este aumento está producido por varios factores, entre los que se encuentran: 1) la apertura de muchos capilares pulmonares previamente cerrados o la dilatación adicional de capilares ya abiertos, aumentando de esta manera el área superficial de la sangre hacia la que puede difundir el O2, y 2) un mejor equilibrio entre la ventilación de los alvéolos y la perfusión de los capilares alveolares con sangre, denominado cociente de ventilación-perfusión
Capacidad de difusión del dióxido de carbono las mediciones de la difusión de otros gases han mostrado que la capacidad de difusión varía directamente con el coeficiente de difusión del gas particular. Como el coeficiente de difusión del CO2 es algo mayor de 20 veces el del O2, cabe esperar que la capacidad de difusión del CO2 en reposo sea de aproximadamente 400 a 450 ml/min/mmHg y durante el esfuerzo de aproximadamente 1.200 a 1.300 ml/min/mmHg
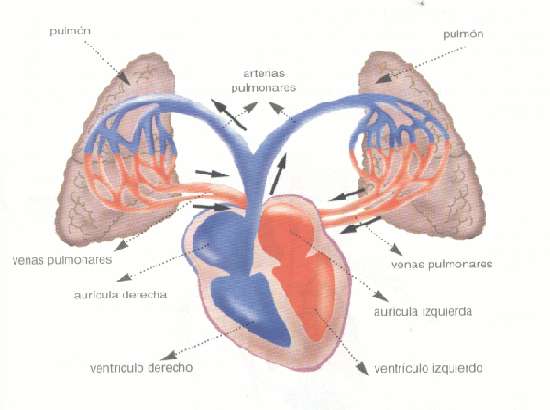
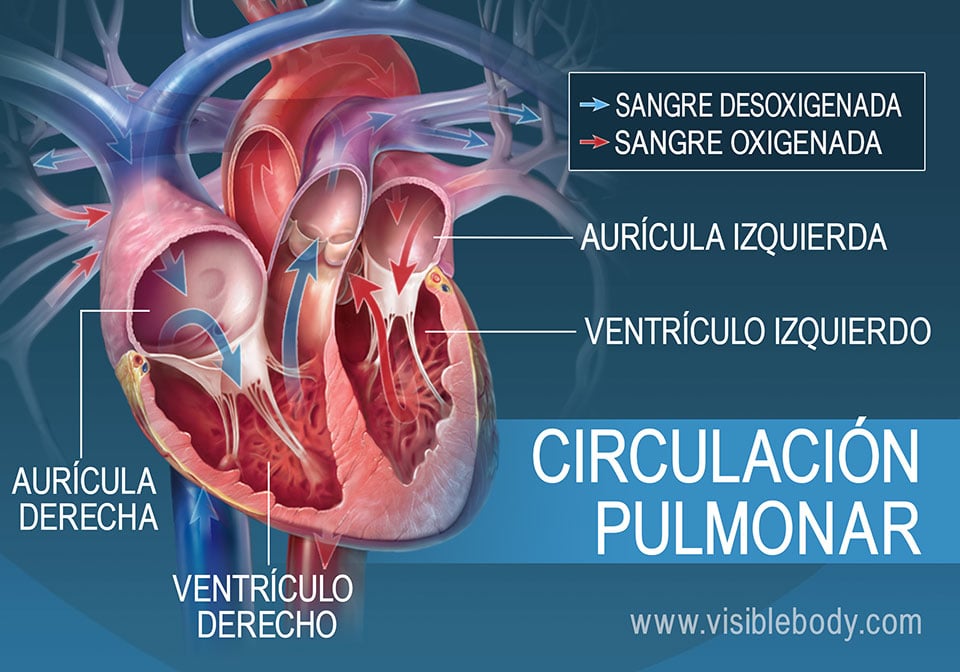
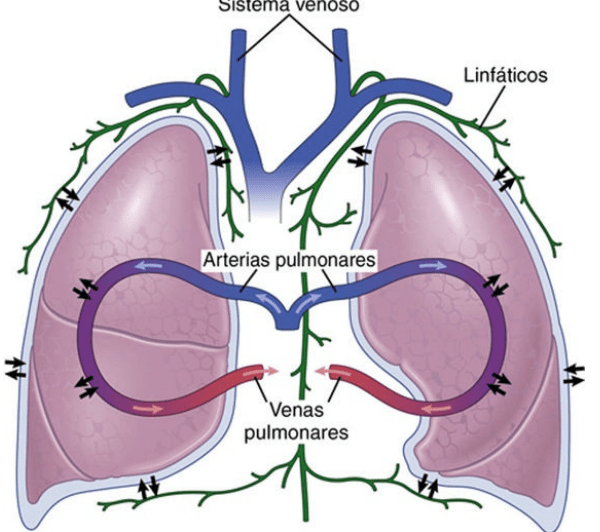
una circulación de bajo flujo y alta presión y una circulación de alto flujo y baja presión. La circulación de bajo flujo y alta presión aporta la sangre arterial sistémica a la tráquea, el árbol bronquial incluidos los bronquíolos terminales, los tejidos de sostén del pulmón y las capas exteriores (adventicias) de las arterias y venas pulmonares. Las arterias bronquiales, que son ramas de la aorta torácica, irrigan la mayoría de esta sangre arterial sistémica a una presión solo inferior a la presión aórtica. La circulación de alto flujo y baja presión que suministra la sangre venosa de todas las partes del organismo a los capilares alveolares en los que se añade el oxígeno (O2) y se extrae el dióxido de carbono (CO2). La arteria pulmonar, que recibe sangre del ventrículo derecho, y sus ramas arteriales transportan sangre a los capilares alveolares para el intercambio gaseoso y a las venas pulmonares y después devuelven la sangre a la aurícula izquierda para su bombeo por el ventrículo izquierdo a través de la circulación sistémica.
Anatomía fisiológica del sistema circulatorio pulmonar
Vasos pulmonares La arteria pulmonar tiene un grosor de pared un tercio del de la aorta. Las ramas de las arterias pulmonares son cortas, y todas las arterias pulmonares, incluso las arterias más pequeñas y las arteriolas, tienen diámetros mayores que sus correspondientes arterias sistémicas. Este aspecto, combinado con el hecho de que los vasos son delgados y distensibles, da al árbol arterial pulmonar una gran distensibilidad, que es en promedio de casi 7 ml/mmHg, que es similar a la de todo el árbol arterial sistémico. Esta gran distensibilidad permite que las arterias pulmonares se acomoden al gasto del volumen sistólico del ventrículo derecho Las venas pulmonares, al igual que las arterias pulmonares, también son cortas. Drenan inmediatamente la sangre que les llega hacia la aurícula izquierda
Vasos bronquiales La sangre también fluye hacia los pulmones a través de arterias bronquiales pequeñas que se originan en la circulación sistémica y transportan el 1-2% del gasto cardíaco total. Esta sangre arterial bronquial es sangre oxigenada, al contrario de la sangre parcialmente desoxigenada de las arterias pulmonares. Vascularizan los tejidos de soporte de los pulmones, como el tejido conjuntivo, los tabiques y los bronquios grandes y pequeños
Linfáticos Hay vasos linfáticos en todos los tejidos de soporte del pulmón, comenzando en los espacios tisulares conjuntivos que rodean a los bronquíolos terminales, y siguiendo hacia el hilio del pulmón, y desde aquí principalmente hacia el conducto linfático torácico derecho. Las sustancias en forma de partículas que entran en los alvéolos son retiradas parcialmente por medio de estos conductos, y también eliminan de los tejidos pulmonares las proteínas plasmáticas que escapan de los capilares pulmonares, contribuyendo de esta manera a prevenir el edema pulmonar
Presiones en el sistema pulmonar
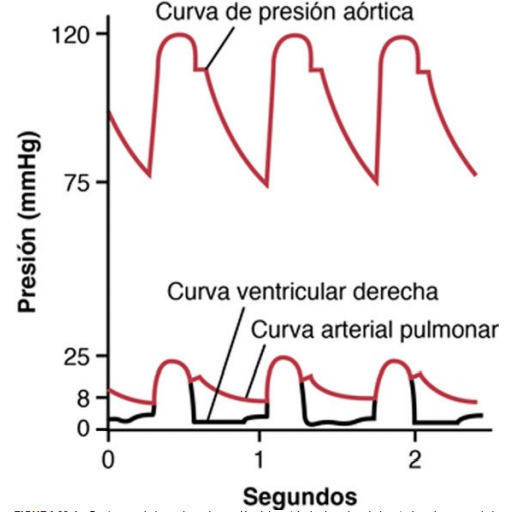
Curva del pulso de presión del ventrículo derecho La presión sistólica del ventrículo derecho del ser humano normal es en promedio de aproximadamente 25 mmHg, y la presión diastólica es en promedio de aproximadamente 0 a 1 mmHg, valores que son solo un quinto de los del ventrículo izquierdo
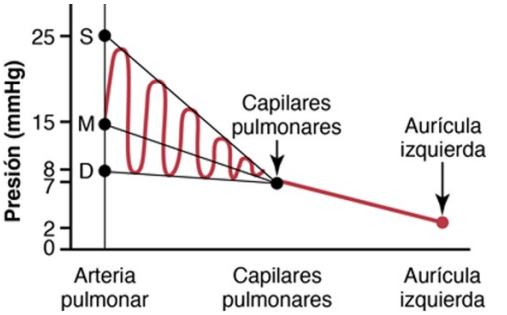
Presiones en la arteria pulmonar la presión arterial pulmonar sistólica se sitúa normalmente en promedio en unos 25 mmHg en el ser humano, la presión arterial pulmonar diastólica es de aproximadamente 8 mmHg y la presión arterial pulmonar media es de 15 mmHg.
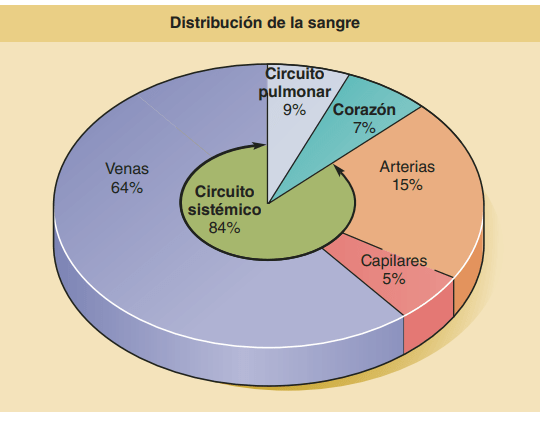
Volumen sanguíneo de los pulmones
El volumen de la sangre de los pulmones es de aproximadamente 450 ml, aproximadamente el 9% del volumen de sangre total de todo el aparato circulatorio. Aproximadamente 70 ml de este volumen de sangre pulmonar están en los capilares pulmonares, y el resto se divide aproximadamente por igual entre las arterias y las venas pulmonares.
La patología cardíaca puede desplazar sangre desde la circulación sistémica a la circulación pulmonar La insuficiencia del lado izquierdo del corazón o el aumento de la resistencia al flujo sanguíneo a través de la válvula mitral como consecuencia de una estenosis mitral o una insuficiencia mitral hace que la sangre quede estancada en la circulación pulmonar, aumentando a veces el volumen de sangre pulmonar hasta un 100% y produciendo grandes aumentos de las presiones vasculares pulmonares. Dado que el volumen de la circulación sistémica es aproximadamente nueve veces el de la circulación pulmonar, el desplazamiento de sangre desde un sistema hacia el otro afecta mucho al sistema pulmonar, pero habitualmente tiene solo efectos circulatorios sistémicos leve
Flujo sanguíneo a través de los pulmones y su distribución
El flujo sanguíneo a través de los pulmones es esencialmente igual al gasto cardíaco. Por tanto, los factores que controlan el gasto cardíaco (principalmente factores periféricos, también controlan el flujo sanguíneo pulmonar. En la mayoría de las situaciones los vasos pulmonares actúan como tubos distensibles que se dilatan al aumentar la presión y se estrechan al disminuir la presión. Para que se produzca una aireación adecuada de la sangre, esta debe distribuirse a los segmentos de los pulmones en los que los alvéolos estén mejor oxigenados. Esta distribución se consigue por el mecanismo siguiente.
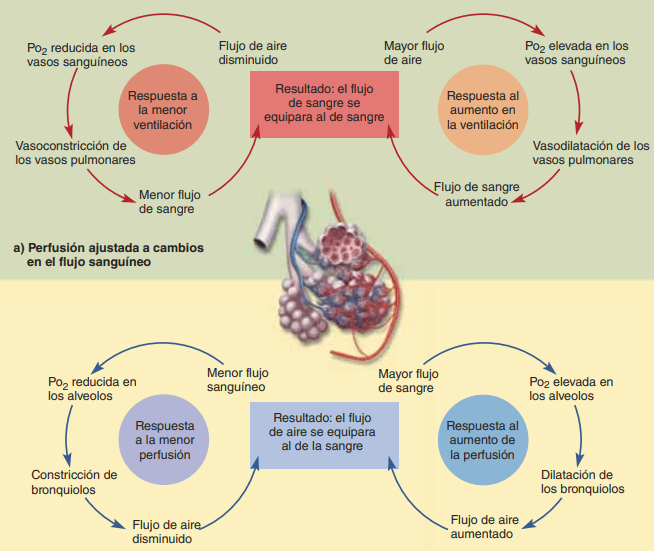
La disminución del oxígeno alveolar reduce el flujo sanguíneo alveolar local y regula la distribución del flujo sanguíneo pulmonar Cuando la concentración de O2 en el aire de los alvéolos disminuye por debajo de lo normal (especialmente cuando disminuye por debajo del 70% de lo normal [es decir, por debajo de 73 mmHg de Po2 ]) los vasos sanguíneos adyacentes se constriñen, con un aumento de la resistencia vascular de más de cinco veces a concentraciones de O2 muy bajas. Este efecto es opuesto al efecto que se observa en los vasos sistémicos, que se dilatan en lugar de constreñirse en respuesta a concentraciones bajas de O2
El aumento en la resistencia vascular pulmonar como consecuencia de una baja concentración de O2 tiene una función importante de distribución del flujo sanguíneo allí donde sea más eficaz. Es decir, si algunos alvéolos están mal ventilados y tienen una concentración baja de O2 , los vasos locales se constriñen. Esta constricción hace que la sangre fluya a través de otras zonas de los pulmones que están mejor aireadas, proporcionando de esta manera un sistema de control automático para distribuir el flujo sanguíneo a las zonas pulmonares en proporción a sus presiones alveolares de oxígeno
Efecto de los gradientes de presión hidrostática de los pulmones sobre el flujo sanguíneo pulmonar regional
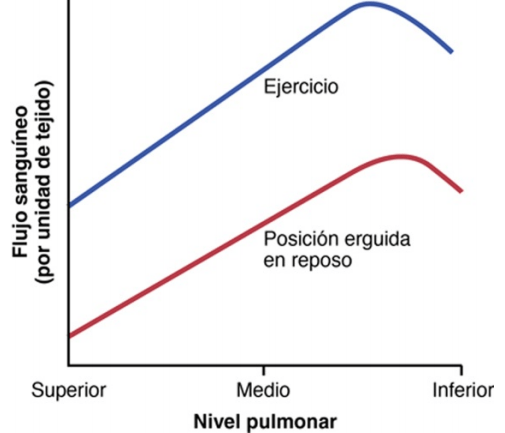
En el adulto en posición erguida el punto más bajo de los pulmones está normalmente unos 30 cm por debajo del punto más alto, lo que representa una diferencia de presión de 23 mmHg, de los cuales aproximadamente 15 mmHg están por encima del corazón y 8 por debajo. Es decir, la presión arterial pulmonar en la porción más elevada del pulmón de una persona que está de pie es aproximadamente 15 mmHg menor que la presión arterial pulmonar a nivel del corazón, y la presión en la porción más inferior de los pulmones es aproximadamente 8 mmHg mayor.
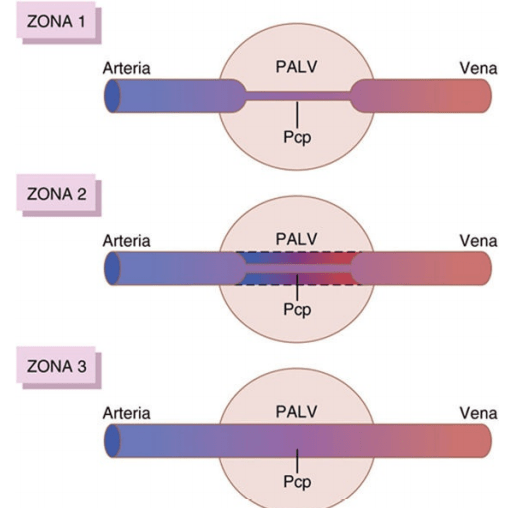
Zonas 1, 2 y 3 del flujo sanguíneo pulmonar Los capilares de las paredes alveolares están distendidos por la presión de la sangre que hay en su interior, pero simultáneamente están comprimidos por la presión del aire alveolar que está en su exterior. Por tanto, siempre que la presión del aire alveolar pulmonar sea mayor que la presión de la sangre capilar, los capilares se cierran y no hay flujo sanguíneo. En diferentes situaciones normales y patológicas se puede encontrar una cualquiera de tres posibles zonas (patrones) del flujo sanguíneo pulmonar: Zona 1: ausencia de flujo durante todas las porciones del ciclo cardíaco porque la presión capilar alveolar local en esa zona del pulmón nunca aumenta por encima de la presión del aire alveolar en ninguna fase del ciclo cardíaco. Zona 2: flujo sanguíneo intermitente, solo durante los picos de presión arterial pulmonar, porque la presión sistólica en ese momento es mayor que la presión del aire alveolar, pero la presión diastólica es menor que la presión del aire alveolar. Zona 3: flujo de sangre continuo, porque la presión capilar alveolar es mayor que la presión del aire alveolar durante todo el ciclo cardíaco
Función de la circulación pulmonar cuando la presión auricular izquierda se eleva como consecuencia de una insuficiencia cardíaca izquierda
cuando se produce insuficiencia del lado izquierdo del corazón la sangre comienza a acumularse en la aurícula izquierda. Como consecuencia, la presión auricular izquierda puede aumentar de manera ocasional desde su valor normal de 1 a 5 mmHg hasta 40 a 50 mmHg. La elevación inicial de la presión auricular, de hasta aproximadamente 7 mmHg, tiene poco efecto sobre la función de la circulación pulmonar. Sin embargo, cuando la presión auricular izquierda aumenta a más de 7 u 8 mmHg, aumentos adicionales de la presión auricular izquierda producen aumentos casi igual de grandes de la presión arterial pulmonar, generando de esta manera un aumento asociado de la carga del corazón derecho. Cualquier aumento de la presión auricular izquierda por encima de 7 u 8 mmHg aumenta la presión capilar casi en la misma magnitud.
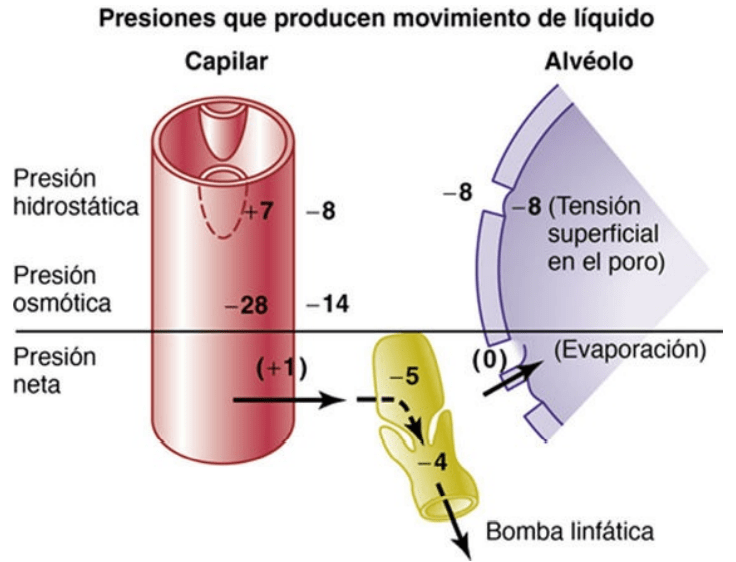
Intercambio capilar de líquido en los pulmones y dinámica del líquido intersticial pulmonar
La presión capilar pulmonar es baja, de aproximadamente 7 mmHg, en comparación con una presión capilar funcional mucho mayor en los tejidos periféricos, de aproximadamente 17 mmHg.
La presión del líquido intersticial del pulmón es ligeramente más negativa que en el tejido subcutáneo periférico. (Esta presión se ha medido de dos formas: con una micropipeta insertada en el intersticio pulmonar, que da un valor de aproximadamente –5 mmHg, y midiendo la presión de absorción de líquido desde los alvéolos, que da un valor de aproximadamente –8 mmHg.)
La presión coloidosmótica del líquido intersticial pulmonar es de aproximadamente 14 mmHg, en comparación con menos de la mitad de este valor en los tejidos periféricos.
Las paredes alveolares son muy delgadas, y el epitelio alveolar que recubre las superficies alveolares es tan débil que se puede romper si la presión positiva en los espacios intersticiales es mayor que la presión del aire alveolar (>0 mmHg), lo que permite el paso de líquido desde los espacios intersticiales hacia los alvéolos.
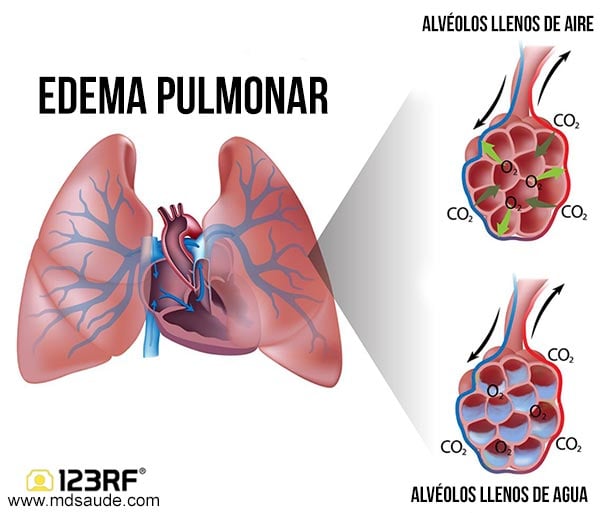
Edema pulmonar
Cualquier factor que aumente la filtración de líquido fuera de los capilares pulmonares o que impida la función linfática pulmonar y provoque un aumento de la presión del líquido intersticial pulmonar desde el intervalo negativo hasta el intervalo positivo dará lugar al llenado rápido de los espacios intersticiales pulmonares y de los alvéolos con grandes cantidades de líquido libre. Las causas más frecuentes de edema pulmonar son:
Insuficiencia cardíaca izquierda o valvulopatía mitral, con los consiguientes grandes aumentos de la presión venosa pulmonar y de la presión capilar pulmonar y el encharcamiento de los espacios intersticiales y de los alvéolos.
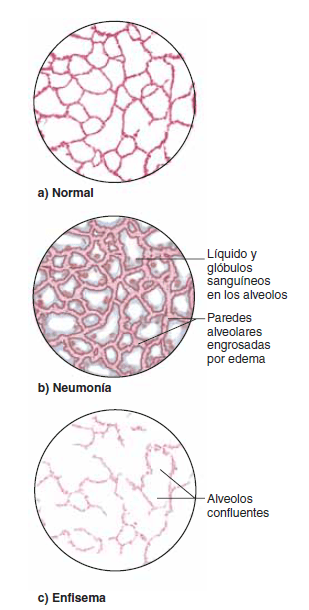
La lesión de las membranas de los capilares sanguíneos pulmonares producida por infecciones como la neumonía o por la inhalación de sustancias tóxicas como el gas cloro o el gas dióxido de azufre. Cada uno de estos mecanismos da lugar a una fuga rápida tanto de proteínas plasmáticas como de líquido desde los capilares hacia los espacios intersticiales pulmonares y los alvéolos.
Líquido en la cavidad pleural
Cuando los pulmones se expanden y se contraen durante la respiración normal se deslizan en el interior de la cavidad pleural. Para facilitar este movimiento hay una delgada capa de líquido mucoide entre las pleuras parietal y visceral. La membrana pleural es una membrana serosa mesenquimatosa porosa a través de la cual trasudan continuamente pequeñas cantidades de líquido intersticial hacia el espacio pleural. Estos líquidos arrastran con ellos proteínas tisulares, lo que da al líquido pleural una característica mucoide, que es lo que permite el deslizamiento muy fácil de los pulmones en movimiento
La cantidad total de líquido en cada una de las cavidades pleurales normalmente es pequeña, solo de algunos mililitros. Siempre que la cantidad sea superior a la justa para comenzar a fluir en la cavidad pleural, el exceso de líquido es extraído mediante bombeo por los vasos linfáticos
Derrame pleural: acumulación de grandes cantidades de líquido libre en el espacio pleural
Las causas del derrame son las mismas que las causas del edema en otros tejidos, entre ellas: 1) bloqueo del drenaje linfático desde la cavidad pleural; 2) insuficiencia cardíaca, que da lugar a unas presiones capilares periférica y pulmonar excesivamente altas, que dan lugar a una trasudación excesiva de líquido hacia la cavidad pleural; 3) marcada reducción de la presión osmótica coloidal del plasma, que permite una trasudación excesiva de líquidos, y 4) infección o cualquier otra causa de inflamación de las superficies de la cavidad pleural, que aumenta la permeabilidad de las membranas capilares y permite la salida rápida tanto de proteínas plasmáticas como de líquido hacia la cavidad.
Los cuatro componentes principales de la respiración son:
1) ventilación pulmonar, que se refiere al flujo de entrada y salida de aire entre la atmósfera y los alvéolos pulmonares 2) difusión de oxígeno (O2) y de dióxido de carbono (CO2) entre los alvéolos y la sangre; 3) transporte de oxígeno y de dióxido de carbono en la sangre y los líquidos corporales hacia las células de los tejidos corporales y desde las mismas, y 4) regulación de la ventilación y otras facetas de la respiración
Mecánica de la ventilación pulmonar
Músculos que causan la expansión y contracción pulmonar
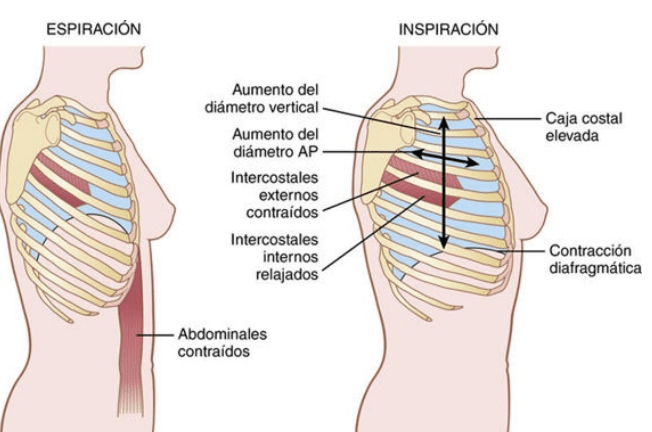
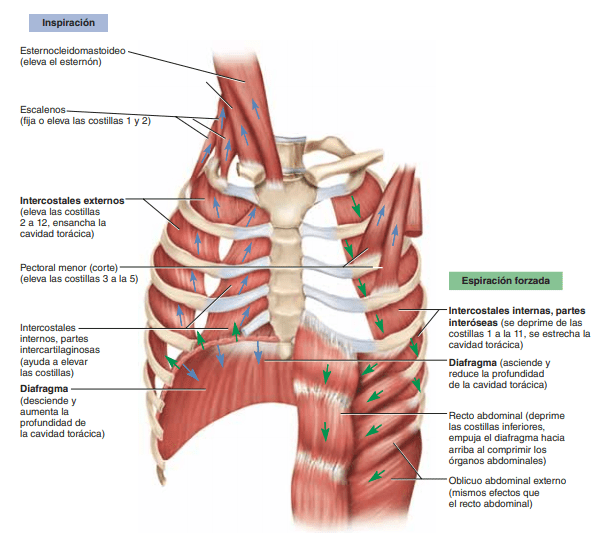
Los pulmones se pueden expandir y contraer de dos maneras: 1) mediante el movimiento hacia abajo y hacia arriba del diafragma para alargar o acortar la cavidad torácica, y 2) mediante la elevación y el descenso de las costillas para aumentar y reducir el diámetro anteroposterior de la cavidad torácica
La respiración tranquila normal se consigue casi totalmente por el primer mecanismo, es decir, por el movimiento del diafragma. Durante la inspiración la contracción del diafragma tira hacia abajo de las superficies inferiores de los pulmones. Después, durante la espiración el diafragma simplemente se relaja, y el retroceso elástico de los pulmones, de la pared torácica y de las estructuras abdominales comprime los pulmones y expulsa el aire. Sin embargo, durante la respiración forzada las fuerzas elásticas no son suficientemente potentes para producir la espiración rápida necesaria, de modo que se consigue una fuerza adicional principalmente mediante la contracción de los músculos abdominales, que empujan el contenido abdominal hacia arriba contra la parte inferior del diafragma, comprimiendo de esta manera los pulmones.
Los músculos más importantes que elevan la caja torácica son los intercostales externos, aunque otros músculos que contribuyen son: 1) los músculos esternocleidomastoideos, que elevan el esternón; 2) los serratos anteriores, que elevan muchas de las costillas, y 3) los escalenos, que elevan las dos primeras costillas.
Los músculos que tiran hacia abajo de la caja costal durante la espiración son principalmente 1) los rectos del abdomen, que tienen el potente efecto de empujar hacia abajo las costillas inferiores al mismo tiempo que ellos y otros músculos abdominales también comprimen el contenido abdominal hacia arriba contra el diafragma, y 2) los intercostales internos.
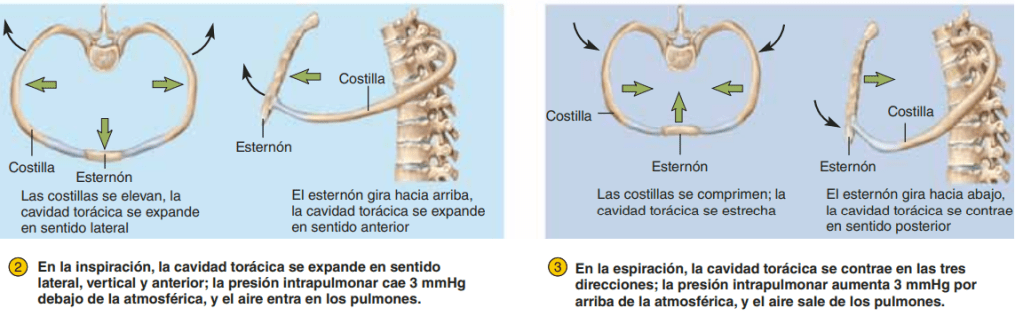
Presiones que originan el movimiento de entrada y salida de aire de los pulmones
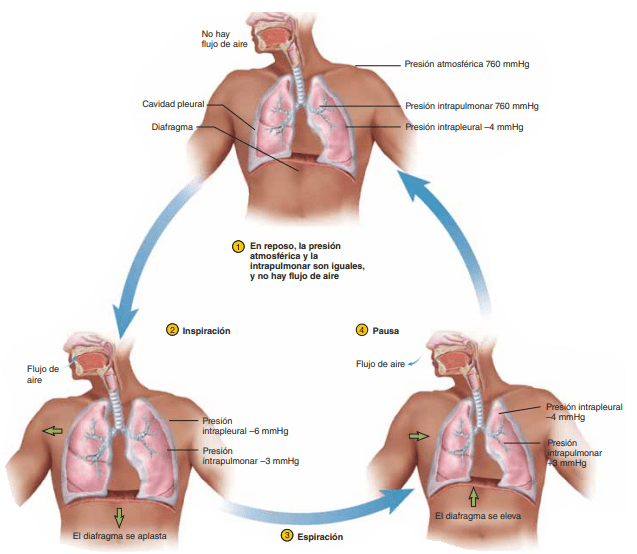
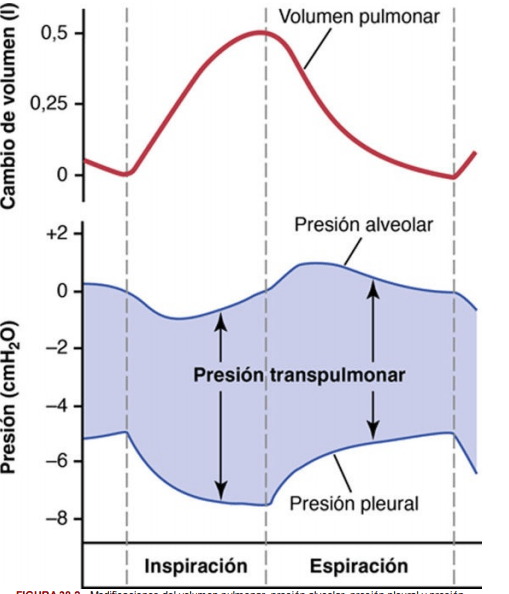
Presión pleural y sus cambios durante la respiración
La presión pleural es la presión del líquido que está en el delgado espacio que hay entre la pleura pulmonar y la pleura de la pared torácica. Como se ha señalado antes, esta presión es normalmente una aspiración ligera, lo que significa que hay una presión ligeramente negativa. La presión pleural normal al comienzo de la inspiración es de aproximadamente –5 cmH2O, que es la magnitud de la aspiración necesaria para mantener los pulmones expandidos hasta su nivel de reposo. Durante la inspiración normal, la expansión de la caja torácica tira hacia fuera de los pulmones con más fuerza y genera una presión más negativa, hasta un promedio de aproximadamente –7,5 cmH2O.
Presión alveolar: presión del aire en el interior de los alvéolos pulmonares
Cuando la glotis está abierta y no hay flujo de aire hacia el interior ni el exterior de los pulmones, las presiones en todas las partes del árbol respiratorio, hasta los alvéolos, son iguales a la presión atmosférica, que se considera que es la presión de referencia cero en las vías aéreas (es decir, presión de 0 cmH2O). Para que se produzca un movimiento de entrada de aire hacia los alvéolos durante la inspiración, la presión en los alvéolos debe disminuir hasta un valor ligeramente inferior a la presión atmosférica (debajo de cero).
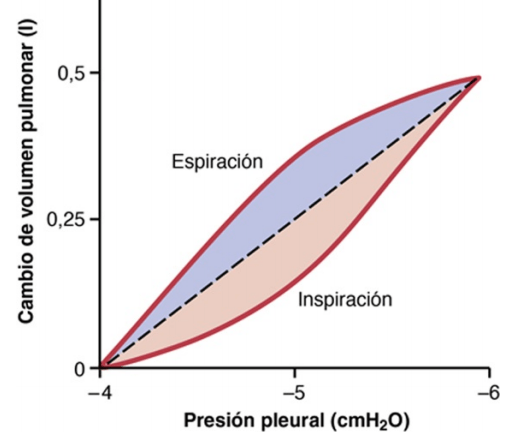
Diagrama de distensibilidad de los pulmones
un diagrama que relaciona los cambios del volumen pulmonar con los cambios de la presión pleural, lo que, a su vez, modifica la presión transpulmonar. que la relación es diferente para la inspiración y para la espiración. Cada una de las curvas se registra modificando la presión pleural en escalones pequeños y permitiendo que el volumen pulmonar llegue a un nivel estable entre escalones sucesivos. Las dos curvas se denominan, respectivamente, la curva de distensibilidad inspiratoria y la curva de distensibilidad espiratoria y todo el diagrama se denomina diagrama de distensibilidad de los pulmones.
Surfactante, tensión superficial y colapso de los alvéolos
Principio de la tensión superficial Cuando el agua forma una superficie con el aire, las moléculas de agua de la superficie del agua tienen una atracción especialmente intensa entre sí. En consecuencia, la superficie del agua siempre está intentando contraerse. Esto es lo que mantiene unidas entre sí las gotas de lluvia: una membrana muy contráctil de moléculas de agua que rodea toda la superficie de la gota de agua. Invirtamos ahora estos principios y veamos qué ocurre en las superficies internas de los alvéolos. Aquí la superficie de agua también intenta contraerse, lo que tiende a expulsar el aire de los alvéolos a través de los bronquios y, al hacerlo, hace que los alvéolos intenten colapsarse. El efecto neto es producir una fuerza contráctil elástica de todo el pulmón, que se denomina fuerza elástica de la tensión superficial
El surfactante es una mezcla compleja de varios fosfolípidos, proteínas e iones. Los componentes más importantes son el fosfolípido dipalmitoilfosfatidilcolina, las apoproteínas del surfactante e iones calcio. La dipalmitoilfosfatidilcolina, junto a otros fosfolípidos menos importantes, es responsable de la reducción de la tensión superficial. Realiza esta función porque no se disuelve de manera uniforme en el líquido que tapiza la superficie alveolar, sino que parte de la molécula se disuelve, mientras que el resto permanece sobre la superficie del agua en los alvéolos. La tensión de esta superficie es entre un doceavo y la mitad de la tensión superficial de una superficie de agua pura.
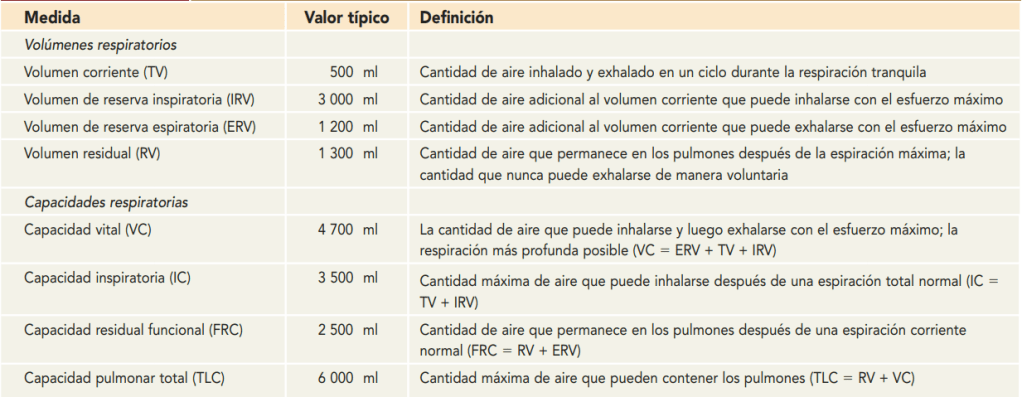
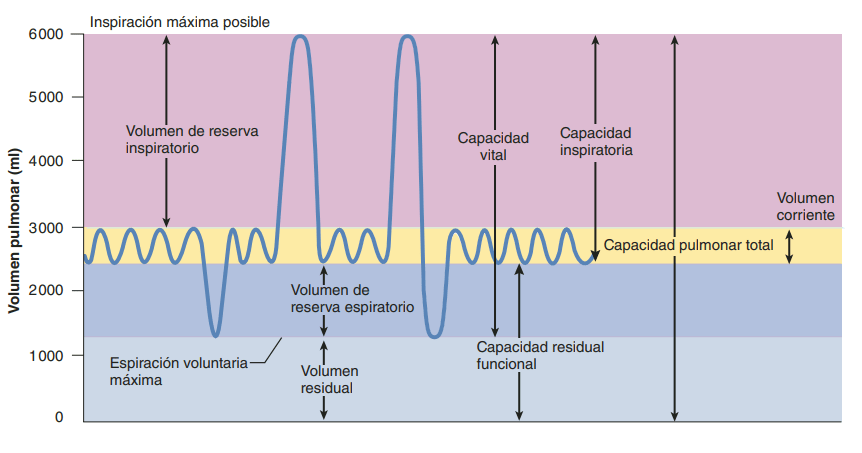
Volúmenes y capacidades pulmonares
Registro de las variaciones del volumen pulmonar: espirometría La ventilación pulmonar puede estudiarse registrando el movimiento del volumen del aire que entra y sale de los pulmones, un método que se denomina espirometría
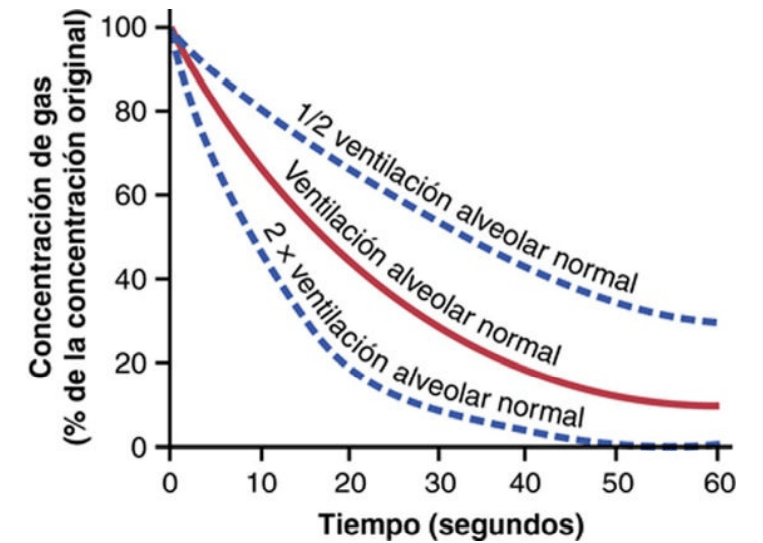
Ventilación alveolar
En último término, la función de la ventilación pulmonar es renovar continuamente el aire de las zonas de intercambio gaseoso de los pulmones, en las que el aire está próximo a la sangre pulmonar. Estas zonas incluyen los alvéolos, los sacos alveolares, los conductos alveolares y los bronquíolos respiratorios. La velocidad a la que llega a estas zonas el aire nuevo se denomina ventilación alveolar
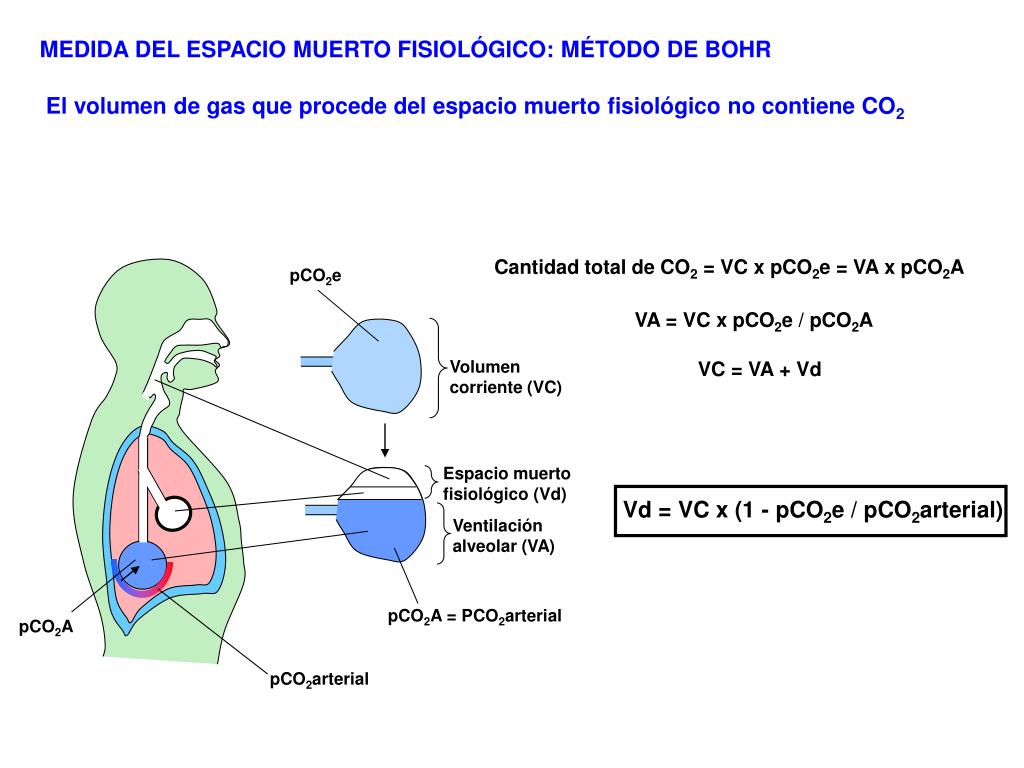
Espacio muerto y su efecto sobre la ventilación alveolar
Parte del aire que respira una persona nunca llega a las zonas de intercambio gaseoso, sino que simplemente llena las vías aéreas en las que no se produce intercambio gaseoso, como la nariz, la faringe la tráquea. Este aire se denomina aire del espacio muerto, porque no es útil para el intercambio gaseoso. Durante la espiración se expulsa primero el aire del espacio muerto, antes de que el aire procedente de los alvéolos llegue a la atmósfera. Por tanto, el espacio muerto es muy desventajoso para retirar los gases espiratorios de los pulmones.
Funciones de las vías aéreas
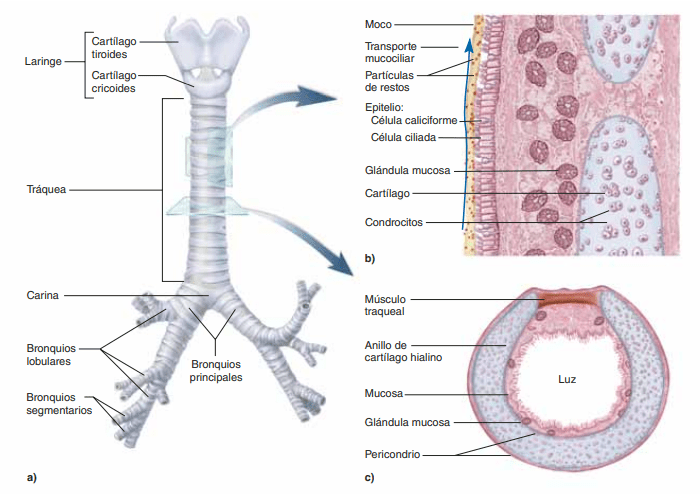
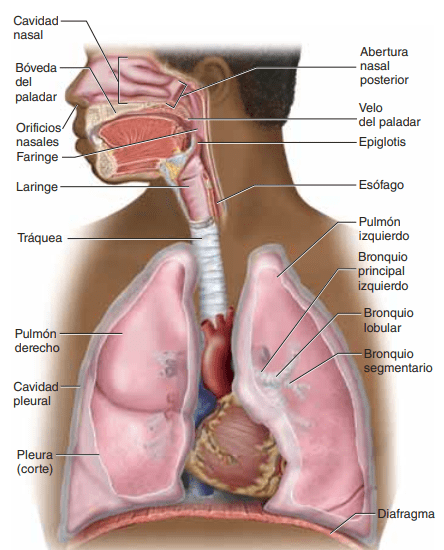
Tráquea, bronquios y bronquíolos El aire se distribuye a los pulmones por medio de la tráquea, los bronquios y los bronquíolos.
Uno de los desafíos más importantes en todas las vías aéreas es mantenerlas abiertas y permitir el paso sin interrupciones de aire hacia los alvéolos y desde los mismos. Para evitar que la tráquea se colapse, múltiples anillos cartilaginosos se extienden aproximadamente 5/6 del contorno de la tráquea. En las paredes de los bronquios, placas curvas de cartílago menos extensas también mantienen una rigidez razonable, aunque permiten un movimiento suficiente para que los pulmones se expandan y se contraigan. Estas placas se hacen cada vez menos extensas en las últimas generaciones de bronquios y han desaparecido en los bronquíolos, que habitualmente tienen diámetros inferiores a 1,5 mm. No se impide el colapso de los bronquíolos por la rigidez de sus paredes. Por el contrario, se mantienen expandidos principalmente por las mismas presiones transpulmonares que expanden los alvéolos. Es decir, cuando los alvéolos se dilatan, los bronquíolos también se dilatan, aunque no tanto
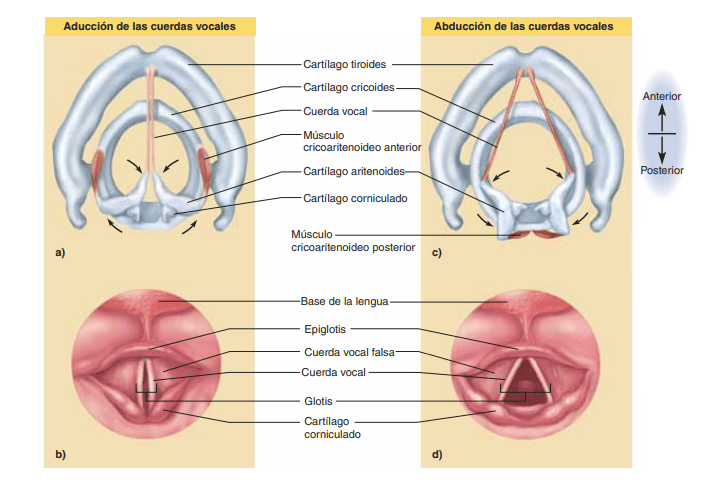
Vocalización El habla implica no solo al aparato respiratorio, sino también a: 1) centros específicos de control nervioso del habla de la corteza cerebral, que se analizan en el capítulo 58; 2) centros de control respiratorio del encéfalo, y 3) las estructuras de articulación y resonancia de las cavidades oral y nasal. El habla está formada por dos funciones mecánicas: 1) fonación, que se realiza en la laringe, y 2) articulación, que se realiza en las estructuras de la boca
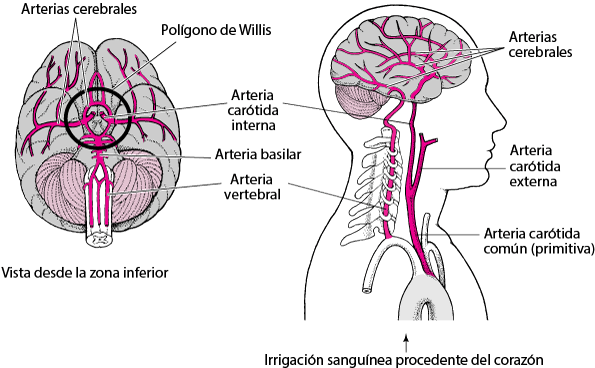
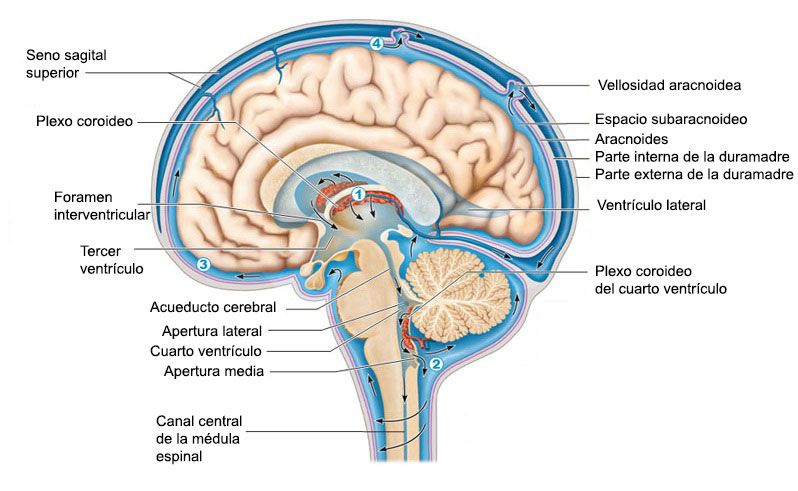
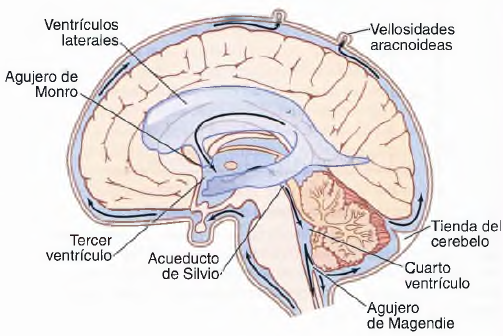
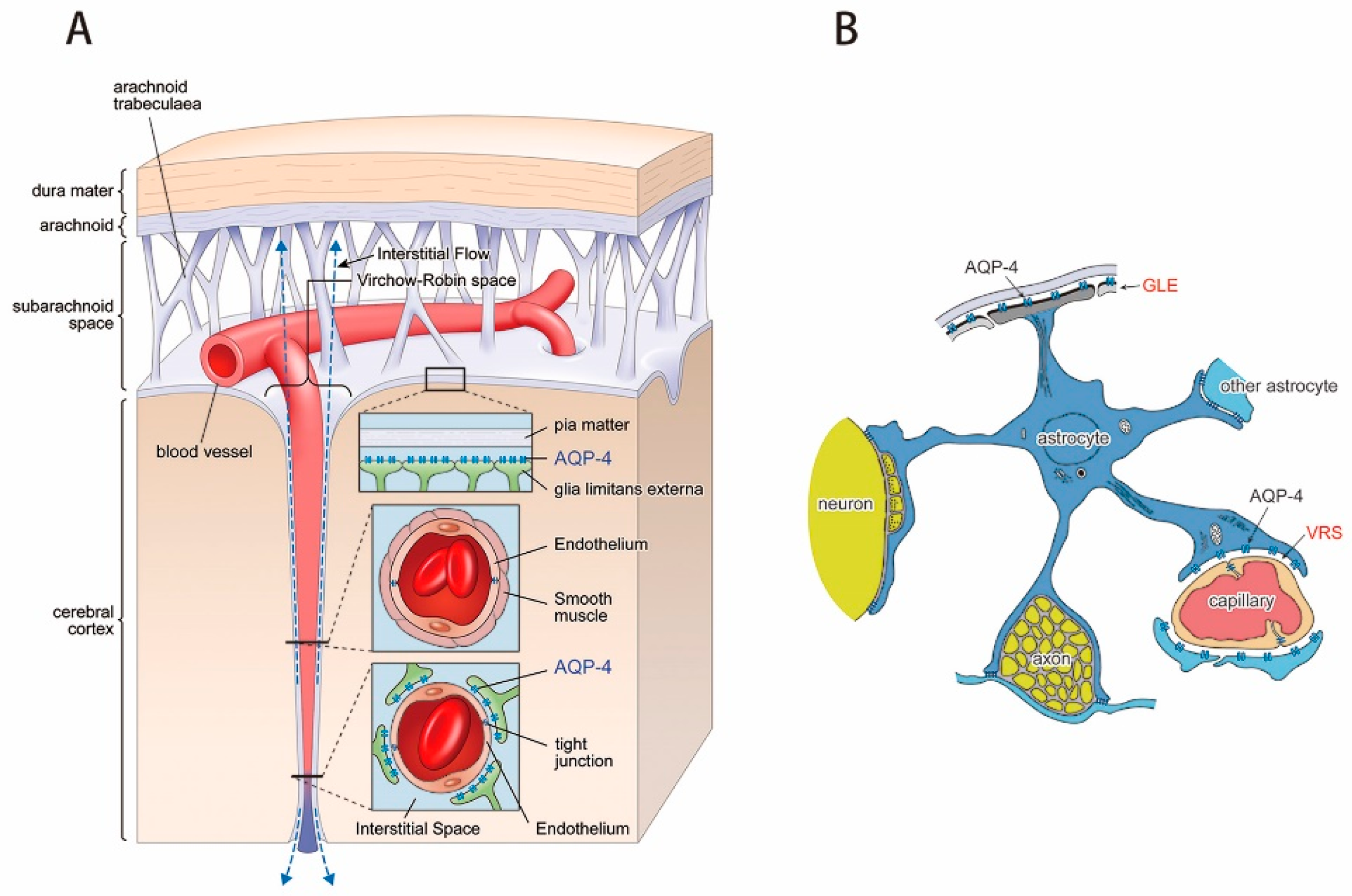
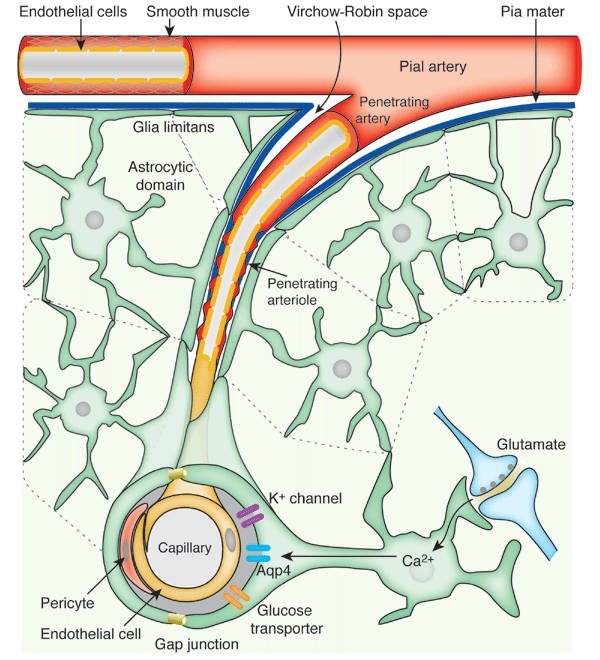
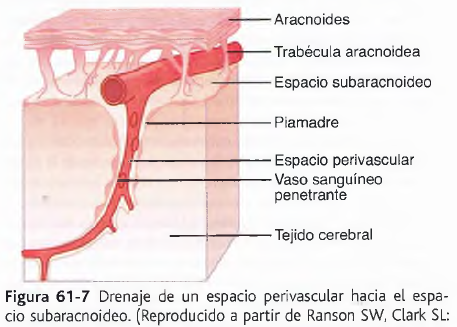
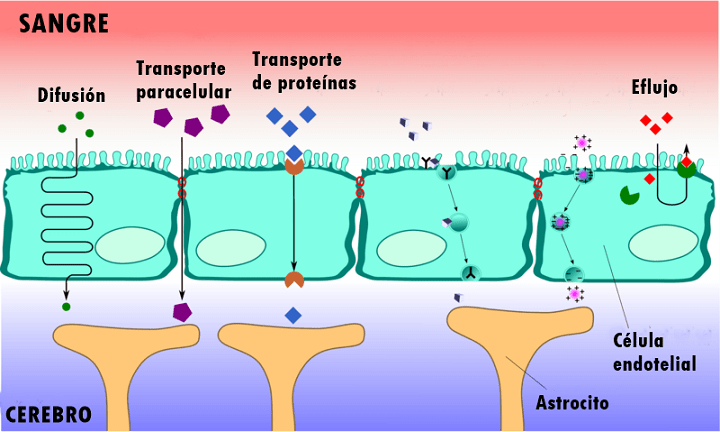
El flujo sanguíneo en el encéfalo es suministrado por cuatro grandes arterias, dos carotídeas y dos vertebrales, que se funden para formar el polígono de Willis en la base del encéfalo. Las arterias que parten del polígono de Willis se desplazan a lo largo de la superficie cerebral y dan origen a las arterias piales, que se ramifican en vasos más pequeños denominados arterias y arteriolas penetrantes. Los vasos penetrantes están separados ligeramente del tejido encefálico por una extensión del espacio subaracnoideo denominada espacio de Virchow-Robin. Los vasos penetrantes se sumergen en el tejido encefálico, para dar lugar a arteriolas intracerebrales, que a su vez se ramifican en capilares en los que tiene lugar el intercambio entre la sangre y los tejidos de oxígeno, nutrientes, dióxido de carbono y metabolitos.
REGULACION DEL FLUJO SANGUÍNEO
Por término medio, el flujo sanguíneo normal a través del cerebro de una persona adulta es de 50 a 65 ml cada 100 g de tejido por minuto. Para todo el encéfalo, esta cantidad asciende 750 a 900 ml/min.Al igual que sucede en la mayoría del resto de tejidos, el flujo sanguíneo cerebral está muy relacionado con el metabolismo tisular. Según se cree, varios factores metabólicos contribuyen a la regulación del flujo sanguíneo cerebral:
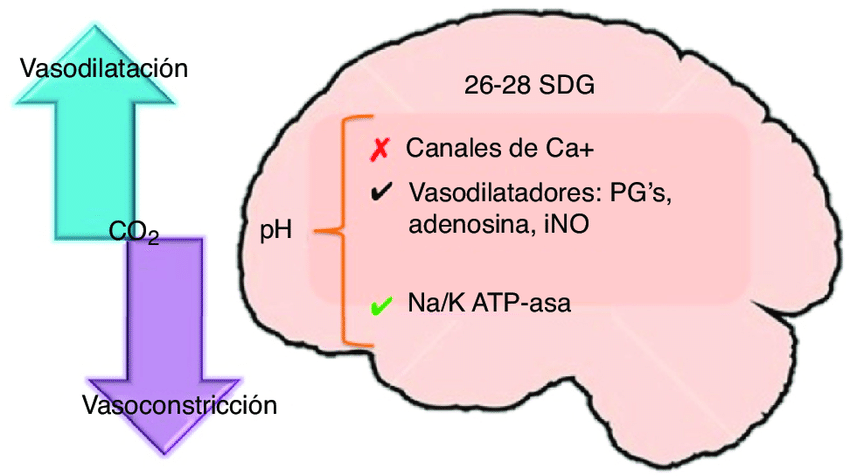
El exceso de concentración de dióxido de carbono o de iones hidrógeno aumenta el flujo sanguíneo cerebral Se cree que el dióxido de carbono incrementa el flujo sanguíneo cerebral al combinarse primero con el agua de los líquidos corporales para formar ácido carbónico, con la posterior disociación de este ácido para producir iones hidrógeno. A continuación, los iones hidrógeno provocan una dilatación de los vasos cerebrales, que es casi directamente proporcional al aumento de su concentración hasta llegar a un límite del flujo sanguíneo, más o menos al doble de lo normal. Otras sustancias que acentuen la acides del tejido cerebral aumentaran el flujo sanguíneo por el mismo procedimiento.
La falta de oxígeno como factor regulador del flujo sanguíneo cerebral Excepto durante los períodos de intensa actividad cerebral, la tasa de utilización del oxígeno por parte del tejido cerebral permanece dentro de unos límites estrechos: es casi exactamente de 3,5 (±0,2) ml de oxígeno cada 100 g de tejido cerebral por minuto. Si, en algún momento, el flujo sanguíneo que llega al encéfalo pasa a ser insuficiente como para suministrar la cantidad necesaria mencionada, la falta de oxígeno causa una vasodilatación casi inmediatamente, con lo que devuelve el flujo sanguíneo cerebral y el transporte de oxígeno hasta los tejidos del cerebro prácticamente a sus condiciones normales.Así pues, este proceso regulador del flujo sanguíneo local es casi exactamente el mismo en el encéfalo que en los vasos sanguíneos coronarios, en el músculo esquelético y en la mayoría de las demás regiones de la circulación corporal.
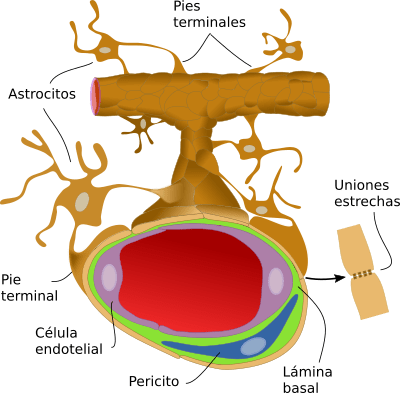
Las sustancias liberadas de los astrocitos regulan el flujo sanguíneo cerebral Los estudios experimentales han demostrado que la estimulación eléctrica de las neuronasglutaminérgicas de excitación conduce a aumentos en la concentración intracelular de iones calcio en lasnprolongaciones alimenticias de los astrocitos y en la dilatación de las arteriolas cercanas. Estudios adicionales han sugerido que la vasodilatación está mediada por varios metabolitos vasoactivos liberados de los astrocitos. Aunque no se sabe claramente cuáles son los mediadores concretos, se ha sugerido que en la mediación de la vasodilatación local son importantes el ácido nítrico, los metabolitos del ácido araquidónico, los iones potasio, la adenosina y otras sustancias generadas por los astrocitos como respuesta a la estimulación de neuronas de excitación adyacentes.
La autorregulación del flujo sanguíneo cerebral protege al cerebrode fluctuaciones en la presión arterial el flujo sanguíneo cerebral está «autorregulado» con suma precisión dentro del intervalo de presión arterial desde 60 hasta 140 mmHg. Es decir, la presión arterial media puede bajar bruscamente hasta 60 mmHg o subir hasta 140 mmHg sin que se produzca ningún cambio apreciable en el flujo que llega. Además, en las personas hipertensas, la autorregulación del flujo sanguíneo cerebral actúa incluso cuando la presión arterial media sube hasta 160 a 180 mmHg.
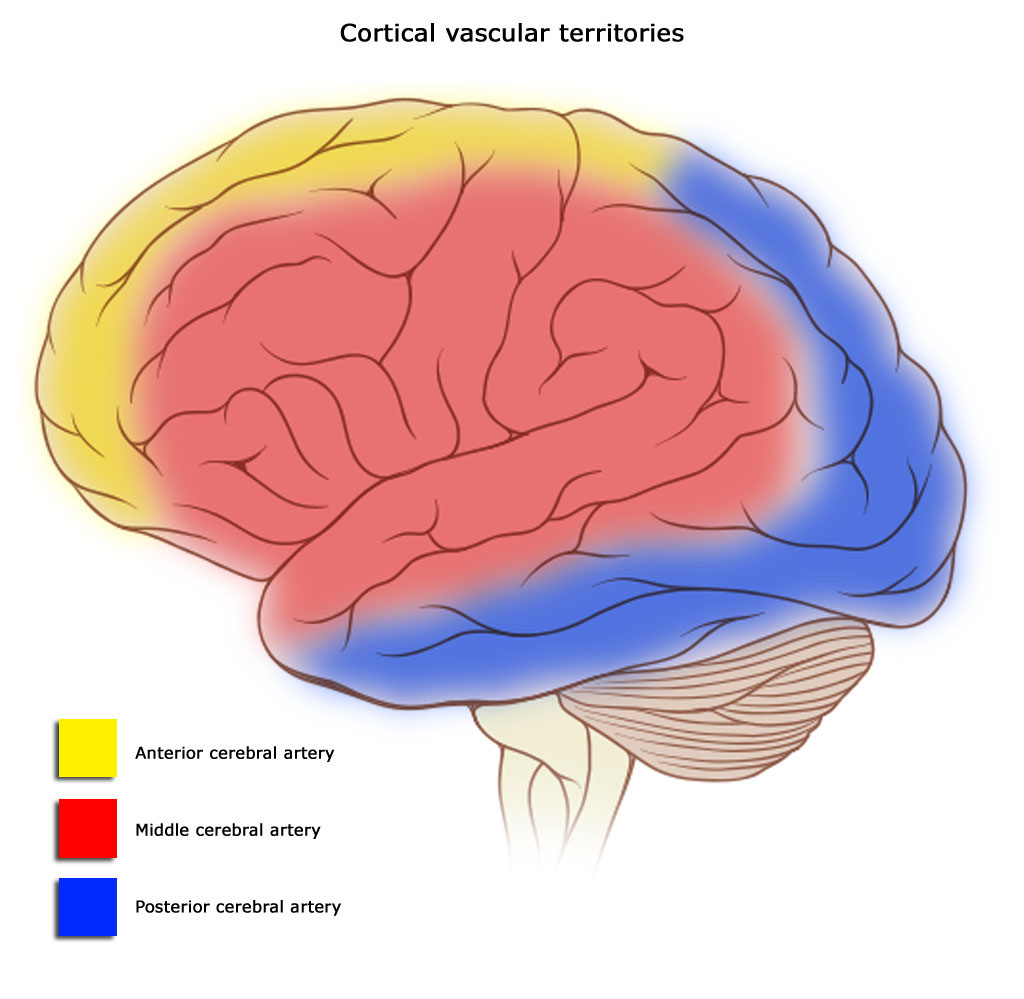
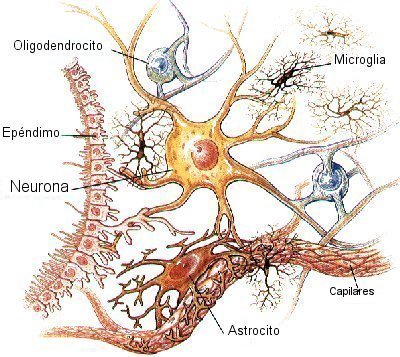
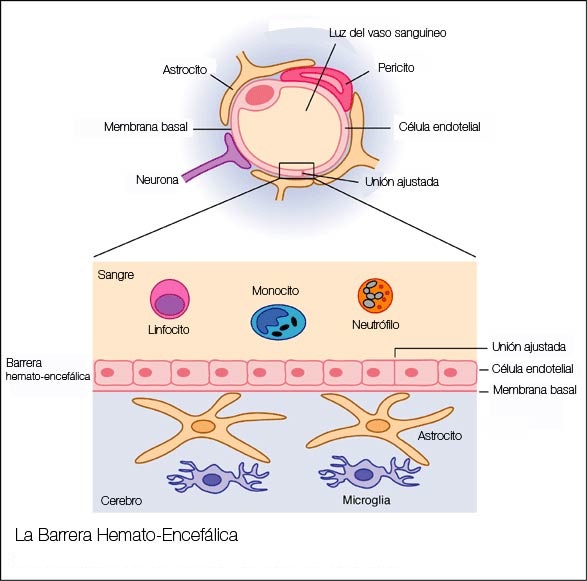
MICROCIRCULACION CEREBRAL Igual que sucede en casi todos los demás tejidos del organismo, el número de capilares sanguíneos en el encéfalo es mayor donde las necesidades metabólicas resulten más grandes. La tasa metabólica global de la sustancia gris cerebral que contiene los somas neuronales es unas cuatro veces mayor que en la sustancia blanca; en consonancia, la cantidad de capilares y la velocidad del flujo sanguíneo también son casi el cuádruple en ella. Una característica estructural importante que presentan los capilares del encéfalo es que en su mayoría son menos «permeables» que los capilares sanguíneos casi de cualquier otro tejido del organismo. Una razón para este fenómeno radica en que cualquiera de sus caras se encuentra reforzada por los «podocitos neurogliales», que consisten en pequeñas prolongaciones procedentes de las células de la glia a su alrededor, que lindan con todas las superficies de los capilares y suministran un soporte físico para impedir su estiramiento excesivo en el caso de que suba demasiado la presión sanguínea capilar. Las paredes de las arteriolas pequeñas que conducen hacia los capilares del encéfalo acaban muy engrosadas en las personas que sufren una elevación de la presión sanguínea, y permanecen notablemente contraídas todo el tiempo para impedir que esta situación se transmita a los capilares
El «ictus» cerebral aparece cuando se obstruyen los vasos sanguíneos cerebrales Casi todos los ancianos tienen bloqueadas algunas arterias pequeñas del encéfalo y hasta el 10% acaba sufriendo a la larga un bloqueo suficiente como para ocasionar un trastorno grave del funcionamiento cerebral, proceso llamado «ictus». La mayoría de los ictus están causados por placas arterioescleróticas que aparecen en una o más de las arterias que irrigan el encéfalo. Las placas tienen la capacidad de activar el mecanismo de la coagulación sanguínea, haciendo que se forme un coágulo y se bloquee el flujo sanguíneo en la arteria, lo que lleva a la pérdida súbita de las funciones cerebrales en un área circunscrita. Más o menos en la cuarta parte de las personas que sufren un ictus, la presión arterial elevada hace que se rompa uno de los vasos sanguíneos; a continuación, se produce una hemorragia, que comprime el tejido cerebral local y altera así su funcionamiento. Los efectos neurológicos de un ictus vienen determinados por la zona afectada del encéfalo
SISTEMA DEL LIQUIDO CEFALORRAQUIDEO Toda la cavidad que encierra el encéfalo y la médula espinal tiene una capacidad de unos 1.600 a 1.700 ml. De ellos, más o menos 150 ml están ocupados por el líquido cefalorraquídeo, y el resto por el encéfalo y la médula. Está presente en los ventrículos cerebrales, en las cisternas que rodean por fuera al encéfalo y en el espacio subaracnoideo alrededor del encéfalo y de la médula espinal. Todas estas cavidades se encuentran conectadas entre sí y la presión del líquido se mantiene a un nivel sorprendentemente constante.
Función amortiguadora del líquido cefalorraquídeo Una función fundamental del líquido cefalorraquídeo consiste en amortiguar el encéfalo dentro de su bóveda sólida. El encéfalo y el líquido cefalorraquídeo poseen aproximadamente la misma densidad específica (tan solo difieren en un 4% más o menos), de modo que el encéfalo se limita a flotar en el seno del líquido. Por tanto, un golpe en la cabeza, si no es demasiado fuerte, desplaza todo el encéfalo a la vez.
Contragolpe Cuando el golpe en la cabeza es intensísimo, puede no dañar el encéfalo en el mismo lado de su acción, aunque es probable que lo dañe en el lado opuesto. Este fenómeno se conoce como «contragolpe» y la razón del mismo es la siguiente: cuando impacta el golpe, el líquido del lado afectado resulta tan incompresible que, al moverse el cráneo, empuja simultáneamente el encéfalo. En el lado opuesto a la zona golpeada, el desplazamiento súbito de todo el cráneo hace que este último se separe transitoriamente con respecto al encéfalo debido a la inercia del cerebro, lo que por un instante crea un espacio vacío en la bóveda craneal del lado opuesto al golpe. Después, cuando el cráneo pierde su aceleración a causa del golpe, el vacío se colapsa bruscamente y el encéfalo choca contra la cara interna del cráneo Si la contusión sucede en el mismo lado donde actúa el impacto, es una lesión por golpe; si ocurre en el lado opuesto, es una lesión por contragolpe.
FORMACION, FLUJO Y ABSORCIÓN DEL LIQUIDO CEFALORRAQUIDEO El líquido cefalorraquídeo se forma a una velocidad de unos 500 ml diarios, lo que supone el triple o el cuádruple de su volumen total en todo el sistema. Alrededor de dos tercios o más de esta cantidad se debe a la secreción desde los plexos coroideos en los cuatro ventrículos, sobre todo en los dos ventrículos laterales. Un poco más se produce en la superficie ependimaria de todos los ventrículos y en la aracnoides. Un pequeño porcentaje procede del encéfalo a través de los espacios perivasculares que quedan alrededor de los vasos sanguíneos que atraviesan el encéfalo
los principales canales para el líquido nacen en los plexos coroideos y después siguen el sistema del líquido cefalorraquídeo. La parte segregada en los ventrículos laterales pasa primero hacia el tercer ventrículo; después, tras la incorporación de una mínima cantidad más en esta cavidad, desciende a lo largo del acueducto de Silvio hacia el cuarto ventrículo, donde aún se añade otra minúscula proporción de líquido. Finalmente, sale del cuarto ventrículo por tres pequeños orificios, los dos agujeros laterales de Luschka y el agujero central de Magendie, para penetrar en la cisterna magna, un espacio de líquido que queda detrás del bulbo raquídeo y debajo del cerebelo. La cisterna magna se continúa con el espacio subaracnoideo que rodea al encéfalo y la médula espinal en su integridad. Casi todo el líquido cefalorraquídeo asciende a continuación desde la cisterna magna a través de estos espacios subaracnoideos alrededor del cerebro. Desde aquí, penetra por las múltiples vellosidades aracnoideas que sobresalen hacia el gran seno venoso sagital y otros senos venosos cerebrales, y las atraviesa.
Secrecion por los plexos coroideos La secreción de líquido hacia los ventrículos por el plexo coroideo depende sobre todo del transporteactivo de iones sodio a través de las células epiteliales que tapizan su parte externa. A su vez, los iones sodio arrastran también grandes cantidades de iones cloruro debido a que su carga positiva atrae la negativa de estos últimos. Los dos combinados elevan el contenido de cloruro sódico en el líquido cefalorraquídeo, que está dotado de actividad osmótica, por lo que a continuación provoca la ósmosis casi inmediata de agua a través de la membrana, para aportar el líquido de la secreción. Otros procesos de transporte menos importantes desplazan pequeñas cantidades de glucosa hacia el líquido cefalorraquídeo y extraen iones potasio y bicarbonato hacia los capilares desde su interior.
Espacios perivasculares y líquido cefalorraquídeo Las grandes arterias y venas del encéfalo se hallan sobre su superficie, pero su tramo final penetra hacianel interior, y arrastra una capa de piamadre, la membrana que cubre al encéfalo. La piamadre está muy poco adherida a los vasos, por lo que existe un espacio, el espacio perivascular, entre ella y cada vaso. Así pues, los espacios perivasculares siguen a las arterias y las venas hacia el encéfalo hasta llegar a las arteriolas y las vénulas.
Como sucede en cualquier otro punto del cuerpo, una pequeña cantidad de proteínas se filtra desde los capilares del encéfalo hacia sus espacios intersticiales. Dado que en el tejido cerebral no existen auténticos linfáticos, este exceso proteico sale contenido en el líquido a través de los espacios perivasculares hasta los espacios subaracnoideos. Al llegar a ellos, las proteínas pasan a circular con el líquido cefalorraquídeo, para absorberse hacia las grandes venas cerebrales a través de las vellosidades aracnoideas.
PRESION DEL LIQUIDO CEFALORRAQUÍDEO
Regulación de la presión del líquido cefalorraquídeo por las vellosidades aracnoideas La velocidad normal de formación del líquido cefalorraquídeo permanece muy constante, por lo que sus cambios rara vez constituyen un factor que influya en el control de la presión. En cambio, las vellosidades aracnoideas funcionan como «válvulas» que permiten la salida sin problemas del líquido cefalorraquídeo y de su contenido hacia la sangre de los senos venosos mientras que impiden elretroceso de la sangre en un sentido opuesto. Normalmente, esta acción valvular de las vellosidades deja que el líquido cefalorraquídeo comience a fluir hacia la sangre cuando su presión supera en unos 1,5 mmHg a la de la sangre en los senos venosos. Después, si la presión del líquido cefalorraquídeo todavía sube más, las válvulas se abren con mayor amplitud. En condiciones normales, esta variable casi nunca asciende más que unos pocos milímetros de mercurio por encima de la presión en el interior de los senos venosos cerebrales.
En los estados patológicos, a veces las vellosidades quedan bloqueadas por grandes partículas sólidas, por una fibrosis o por un exceso de células sanguíneas que se hayan filtrado hacia el líquido cefalorraquídeo en los casos de una enfermedad cerebral. Tal bloqueo puede elevar la presión del líquido cefalorraquídeo
Medición de la presión del líquido cefalorraquídeo El procedimiento habitual para medir la presión del líquido cefalorraquídeo es sencillo. En primer lugar, la persona se tumba en posición totalmente horizontal sobre un costado para que la presión del líquido en el conducto raquídeo sea idéntica a la que hay en la bóveda craneal. A continuación, se introduce una aguja de punción en la zona lumbar del conducto raquídeo por debajo del extremo inferior de la médula, y se conecta a un tubo vertical de vidrio cuyo extremo superior está abierto al aire. Se deja que el líquido del conducto vertebral suba por el tubo todo lo que pueda. Si asciende hasta una altura de 136 mm por encima del nivel de la aguja, se dice que su valor es de 136 mm de presión de agua o, dividiendo esta cifra entre 13,6, que es la densidad específica del mercurio, de unos 10 mmHg de presión.
BARRERA HEMATOENCEFALICA Y HEMATOCEFALORRAQUIDEA
muchas moléculas grandes apenas consiguen pasar desde la sangre hacia el líquido cefalorraquídeo o hacia los líquidos intersticiales del encéfalo, aunque estas mismas sustancias salen con facilidad hacia los líquidos intersticiales habituales del organismo. Por tanto, se dice que existen barreras, llamadas barrera hematocefalorraquídea y barrera hematoencefálica, que separan la sangre del líquido cefalorraquídeo y del líquido encefálico, respectivamente. Hay barreras en los plexos coroideos y en las membranas de los capilares tisulares prácticamente en cualquier región del parénquima cerebral excepto en algunas zonas del hipotálamo, la glándula pineal y el área postrema, donde las sustancias difunden sin tantos problemas hacia los espacios tisulares.
En general, las barreras hematocefalorraquídea y hematoencefálica son muy permeables al agua, el dióxido de carbono, el oxígeno y la mayoría de las sustancias liposolubles, como el alcohol y los anestésicos; parcialmente permeables a electrólitos, como el sodio, el cloruro y el potasio, y casi totalmente impermeables a las proteínas plasmáticas y a la mayoría de las moléculas orgánicas grandes no liposolubles.La causa de la baja permeabilidad que presentan las barreras hematocefalorraquídea y hematoencefálica radica en el modo como están unidas entre sí las células endoteliales de los capilares en el tejido cerebral, mediante las denominadas uniones intercelulares herméticas o estrechas.
METABOLISMO CEREBRAL
Índice metabólico cerebral total e índice metabólico de las neuronas En condiciones de vigilia en reposo, al metabolismo cerebral le corresponde aproximadamente el 15% del metabolismo total del organismo, aunque su masa no supone más que el 2% de la masa corporal íntegra. Por tanto, en condiciones de reposo, el metabolismo cerebral por unidad de masa tisular es unas 7,5 veces el metabolismo medio que existe fuera de los tejidos del sistema nervioso. La mayor parte de este metabolismo sucede en las neuronas, no en los tejidos gliales de soporte. La principal necesidad metabólica neuronal consiste en bombear iones a través de sus membranas, sobre todo para transportar sodio y calcio al exterior de la membrana neuronal y potasio a su interior.
Demandas especiales de oxígeno por parte del cerebro: ausencia de un metabolismo anaerobio apreciable El encéfalo no es capaz de efectuar un gran metabolismo anaerobio. Una de las razones para ello estriba en el elevado índice metabólico de las neuronas, por lo que la mayor parte de la actividad neuronal depende de la liberación de oxígeno cada segundo desde la sangre. Si se reúnen todos estos factores, puede entenderse por qué la interrupción brusca del flujo sanguíneo hacia el encéfalo o la ausencia total súbita de oxígeno en la sangre pueden provocar la pérdida del conocimiento en un plazo de 5 a 10 s.
En condiciones normales, la mayor parte de la energía cerebral viene suministrada por la glucosa En condiciones normales, casi toda la energía utilizada por las células del encéfalo llega suministrada por la glucosa extraída de la sangre. Como sucede en el caso del oxígeno, la mayor parte de esta glucosa procede de la sangre capilar minuto a minuto y segundo a segundo, pues sus reservas almacenadas normalmente como glucógeno en las neuronas solo llegan a un total de unos 2 min en cualquier momento determinado. Un rasgo especial que caracteriza la liberación de la glucosa hacia las neuronas es que el transporte a través de la membrana celular no depende de la insulina.
El sistema nervioso autónomo es la porción del sistema nervioso que controla la mayoría de las funciones viscerales del cuerpo. Este componente interviene en la regulación de la presión arterial, la motilidad digestiva, las secreciones gastrointestinales, el vaciamiento de la vejiga urinaria, la sudoración, la temperatura corporal y otras muchas actividades. Algunas de ellas se encuentran casi del todo bajo su dominio en algunos casos y solo parcialmente en otros
Organización general del sistema nervioso autónomo El sistema nervioso autónomo se activa sobre todo a partir de centros situados en la médula espinal, el tronco del encéfalo y el hipotálamo. Asimismo, ciertas porciones de la corteza cerebral, sobre todo de la corteza límbica, pueden transmitir señales hacia los centros inferiores e influir de este modo en el control autónomo. El sistema nervioso autónomo también suele operar por medio de reflejos viscerales. Es decir, las señales sensitivas subconscientes procedentes de órganos viscerales pueden llegar a los ganglios autónomos, el tronco del encéfalo o el hipotálamo, y a continuación devolver unas respuestas reflejas subconscientes directamente a los órganos viscerales para controlar su actividad. Las señales autónomas eferentes se transmiten hacia los diversos órganos del cuerpo a través de sus dos componentes principales, denominados sistema nervioso simpático y sistema nervioso parasimpático,
ANATOMIA FISIOLÓGICA DEL SISTEMA NERVIOSO AUTÓNOMO
Las fibras nerviosas simpáticas nacen en la médula espinal junto a los nervios raquídeos entre los segmentos medulares T1 y L2, y pasan primero a la cadena simpática y después a los tejidos y órganos que resultan estimulados por los nervios simpáticos.
Neuronas simpáticas preganglionares y posganglionares Los nervios simpáticos son diferentes de los nervios motores esqueléticos por el hecho siguiente: cada vía simpática que se dirige desde la médula hasta el tejido estimulado está compuesta por dos células, una neurona preganglionar y una neurona posganglionar, a diferencia de la única neurona existente en la vía motora esquelética. El soma celular de cada neurona preganglionar está situado en el asta intermediolateral de la médula espinal; sus fibras van por una raíz ventral de la médula hasta llegar al nervio raquídeo correspondiente,
Nada más salir el nervio raquídeo del conducto raquídeo, las fibras simpáticas preganglionares lo abandonan y se encaminan a través de un ramo comunicante blanco hacia uno de los ganglios de la cadena simpática. Las fibras pueden seguir uno de los tres trayectos siguientes: 1) hacer sinapsis con neuronas simpáticas posganglionares en el ganglio al que llegan; 2) ascender o descender por la cadena y realizar sinapsis en cualquiera de los otros ganglios que la forman, o 3) recorrer una distancia variable a lo largo de la cadena y después irradiar hacia fuera a través de uno de los nervios simpáticos, para acabar haciendo sinapsis en un ganglio simpático periférico.
Distribución segmentaria de las fibras nerviosas simpáticas Las vías simpáticas que nacen en los diversos segmentos de la médula espinal no tienen por qué distribuirse siguiendo la misma porción corporal que las fibras somáticas del nervio raquídeo correspondiente al mismo segmento. En su lugar, las fibras simpáticas del segmento medular T1 en general 1) ascienden por la cadena simpática para acabar en la cabeza; 2) las pertenecientes a T2 terminan en el cuello; 3) las de T3, T4, T5 y T6 lo hacen en el tórax; 4) las de T7, T8, T9, T10 y T11 en el abdomen, y 5) las de T12, L1 y L2 en las piernas. Esta distribución solo es aproximada y los solapamientos resultan abundantes.
ANATOMIA FISIOLÓGICA DEL SISTEMA NERVIOSO PARASIMPATICO El sistema nervioso parasimpático está representado por fibras que salen del sistema nervioso central a través de los pares craneales III, VII, IX y X; otras fibras parasimpáticas distintas abandonan la parte más inferior de la médula espinal por medio del segundo y el tercer nervio raquídeo sacro y, en ocasiones, por los nervios sacros primero y cuarto. En torno al 75% de todas las fibras nerviosas parasimpáticas están en el nervio vago (par craneal X), llegando a todas las regiones torácicas y abdominales del tronco.
Las fibras parasimpáticas del tercer par craneal llegan al esfínter de la pupila y al músculo ciliar del ojo. Las del séptimo par craneal van dirigidas a las glándulas lagrimal, nasal y submandibular, y las del noveno par craneal se distribuyen por la glándula parótida. Las fibras parasimpáticas sacras están en los nervios pélvicos, que atraviesan el plexo sacro formado por nervios raquídeos a cada lado de la médula en los niveles S2 y S3. A continuación se distribuyen por el colon descendente, el recto, la vejiga urinaria y las porciones inferiores de los uréteres.
CARACTERISTICAS BASICAS DEL FUNCIONAMIENTO SIMPATICO Y PARASIMPATICO Fibras colinérgicas y adrenérgicas: secreción de acetilcolina o noradrenalina
todas o prácticamente todas las terminaciones nerviosas finales del sistema parasimpático segregan acetilcolina. Por el contrario, casi todas las terminaciones nerviosas simpáticas segregan noradrenalina, pero unas pocas segregan acetilcolina. Estos neurotransmisores, a su vez, actúan sobre los distintos órganos para generar los efectos simpáticos o parasimpáticos respectivos. Por tanto, a la acetilcolina se la denomina transmisor parasimpático y a la noradrenalina transmisor simpático
MECANISMOS PARA LA SECRECIÓN DE LOS TRANSMISORES Y SU ELIMINACIÓN EN LAS TERMINACIONES POSGANGLIONARES Secreción de acetilcolina y noradrenalina por las terminaciones nerviosas posganglionares Unas cuantas terminaciones nerviosas autónomas posganglionares, sobre todo las de los nervios parasimpáticos, son semejantes a las de la unión neuromuscular esquelética, pero mucho más pequeñas. Sin embargo, muchas de las fibras nerviosas parasimpáticas y casi todas las simpáticas se limitan meramente a rozar las células efectoras de los órganos inervados a su paso por ellos; o, en algunos casos, terminan en el tejido conjuntivo que ocupa un lugar adyacente a las células que vayan a ser activadas. En el punto donde estos filamentos tocan o pasan sobre las células estimuladas o en su proximidad suelen presentar unas dilataciones bulbosas llamadas varicosidades; es en estas varicosidades donde se sintetizan y almacenan las vesículas transmisoras de la acetilcolina o la noradrenalina.
Receptores de los órganos efectores Antes de que la acetilcolina, la noradrenalina o la adrenalina segregadas en una terminación nerviosa autónoma puedan estimular un órgano efector, primero deben unirse a sus receptores específicos en las células correspondientes. El receptor está situado en el exterior de la membrana celular, ligado como un grupo prostético a una molécula proteica que atraviesa toda la membrana celular. La fijación de la sustancia transmisora al receptor provoca un cambio de configuración en la estructura de la molécula proteica. A su vez, por regla general, la molécula modificada excita o inhibe a la célula: 1) causando un cambio en la permeabilidad de la membrana celular frente a uno o más iones, o 2) activando o inactivando una enzima ligada al otro extremo de la proteína receptora donde sobresale hacia el interior de la célula.
Dos tipos principales de receptores para la acetilcolina: receptores muscarínicos y nicotínicos La acetilcolina activa sobre todo dos tipos de receptores, que reciben la denominación de receptores muscarínicos y nicotínicos. La razón de estos nombres radica en que la muscarina, un producto tóxico de las setas, solo activa los receptores muscarínicos y no los nicotínicos, mientras que la nicotina solo activa los nicotínicos. La acetilcolina estimula ambos. Los receptores muscarínicos, que usan proteínas G como mecanismo de señalización, están presentes en todas las células efectoras estimuladas por las neuronas colinérgicas posganglionares del sistema nervioso parasimpático, así como del sistema simpático. Los receptores nicotínicos son canales iónicos activados por ligando que se observan en los ganglios autónomos, a nivel de las sinapsis entre las neuronas preganglionares y las posganglionares de los sistemas simpático y parasimpático
Receptores adrenérgicos: receptores α y β También hay dos clases de receptores adrenérgicos; se denominan receptores α y receptores β. Existen dos tipos principales de receptores α, α1 y α2 , que se unen a diferentes proteínas G. Los receptores β se dividen en receptores β1 , β2 y β3 porque determinados productos químicos no actúan más que sobre alguno de ellos. Los receptores β también utilizan proteínas G para la señalización.
ACCIONES EXCITADORAS E INHIBIDORAS DE LA ESTIMULACION SIMPATICA Y PARASIMPATICA La estimulación simpática origina unos efectos excitadores en algunos órganos, pero inhibidores en otros. Análogamente, la estimulación parasimpática también causa excitación en algunos e inhibición en otros. Asimismo, cuando la estimulación simpática excita un órgano concreto, a veces la estimulación parasimpática lo inhibe, lo que deja de manifiesto que los dos sistemas en ocasiones actúan recíprocamente entre sí. Sin embargo, la mayoría de los órganos están predominantemente controlados por uno u otro de ellos.
Efectos de la estimulación simpática y parasimpática sobre órganos concretos
Ojos Dos funciones oculares están controladas por el sistema nervioso autónomo: 1) la apertura pupilar, y 2) el enfoque del cristalino. La estimulación simpática contrae las fibras meridionales del iris y dilata la pupila, mientras que la activación parasimpática contrae el músculo circular del iris para contraer la pupila. El parasimpático encargado de controlar la pupila experimenta una estimulación refleja cuando llega a los ojos una luz excesiva; este reflejo reduce la apertura pupilar y disminuye la cantidad de luz que alcanza la retina. Por el contrario, el simpático sufre su estimulación durante los períodos de excitación y aumenta la apertura pupilar en tales circunstancias. El enfoque del cristalino está controlado casi en su integridad por el sistema nervioso parasimpático.
Plexo nervioso intraparietal del aparato digestivo El aparato digestivo dispone de su propia colección intrínseca de nervios, denominada plexo intraparietal o sistema nervioso entérico intestinal y situada en las paredes del intestino. Asimismo, la estimulación tanto simpática como parasimpática procedente del encéfalo puede influir sobre la actividad gastrointestinal sobre todo al potenciar o atenuar las acciones específicas llevadas a cabo por el plexo intraparietal digestivo. En general, la estimulación parasimpática aumenta el grado de actividad global en el tubo digestivo al favorecer el peristaltismo y la relajación de los esfínteres, lo que permite un avance rápido de su contenido.
Corazón En general, la estimulación simpática aumenta la actividad global del corazón. Este efecto se produce mediante un incremento en la frecuencia cardíaca y en la fuerza de la contracción. La estimulación parasimpática provoca básicamente los efectos opuestos: descenso de la frecuenciacardíaca y de la fuerza de la contracción.
FUNCION DE LA MEDULA SUPRARENAL La estimulación de la médula suprarrenal por parte de los nervios simpáticos hace que se libere una gran cantidad de adrenalina y noradrenalina a la circulación sanguínea, y estas dos hormonas a su vez se transportan por la sangre hasta todos los tejidos del cuerpo. Como promedio, más o menos el 80% de la secreción corresponde a adrenalina y el 20% a noradrenalina, aunque sus proporciones relativas pueden cambiar considerablemente en diferentes condiciones fisiológicas. La adrenalina y la noradrenalina circulantes ejercen casi las mismas acciones sobre los diversos órganos que las ocasionadas por la estimulación simpática directa, excepto que sus efectos duran de 5 a 10 veces más debido a que estas dos hormonas desaparecen de la sangre con lentitud en un plazo de 2 a 4 min.
«Tono» simpático y parasimpático Normalmente, los sistemas simpático y parasimpático están constantemente activos, y sus tasas basales de funcionamiento se conocen, respectivamente, como tono simpático y tono parasimpático. El valor de este factor reside en permitir que un solo sistema nervioso aumente o disminuya la actividad de un órgano estimulado
REFLEJOS AUTONOMOS
Reflejos autónomos cardiovasculares Varios reflejos del aparato cardiovascular sirven para controlar la presión arterial y la frecuencia cardíaca. En pocas palabras, los receptores para el estiramiento llamados barorreceptores están situados en las paredes de varias arterias importantes, entre ellas especialmente la arteria carótida interna y el cayado de la aorta. Su extensión debido al aumento de la presión transmite señales hacia el tronco del encéfalo, donde inhiben los impulsos simpáticos destinados al corazón y los vasos sanguíneos y excitan el parasimpático; esto permite el descenso de la presión arterial hasta su normalidad.
Reflejos autónomos digestivos La parte superior del tubo digestivo y el recto están controlados sobre todo por reflejos autónomos. Por ejemplo, el olor de un alimento apetitoso o la presencia de comida en la cavidad oral pone en marcha unas señales que van desde la nariz y la boca hasta los núcleos salivales, glosofaríngeo y vagal del tronco del encéfalo. Estos núcleos, a su vez, envían impulsos a través de los nervios parasimpáticoshasta las glándulas secretoras de la boca y del estómago, lo que da lugar a la producción de jugos gástricos a veces incluso antes de que entre la comida en la boca. Cuando las heces llenan el recto en el extremo opuesto del conducto digestivo, los impulsos sensitivos desencadenados por el estiramiento de este órgano se mandan hasta la porción sacra de la médula espinal y el parasimpático sacro devuelve una señal refleja hasta las partes distales del colon; estas señales producen unas potentes contracciones peristálticas que causan la defecación.
RESPUESTA DE ALARMA O DE ESTRES EN EL SISTEMA NERVIOSO SIMPATICO
Cuando una gran porción del sistema nervioso simpático descarga a la vez, esto aumenta por múltiples vías la capacidad del organismo para realizar una actividad muscular vigorosa de muchas formas, ejemplo lo siguiente: Aumento de la glucólisis hepática y muscular.Aumento de la fuerza muscular.Aumento de la actividad mental.
La suma de todos estos efectos permite que una persona realice una actividad física más extenuante de lo que sería posible en otras condiciones. Dado que el estrés mental o físico pueden excitar el sistema simpático, muchas veces se dice que el objetivo de este componente consiste en suministrar una activación suplementaria al cuerpo en los estados de estrés, que se llama respuesta de estrés simpática. La actividad del sistema simpático adquiere una especial intensidad en muchas situaciones emocionales
CONTROL BULBAR, PONTINO Y MESENCEFALICO DEL SISTEMA NERVIOSO AUTÓNOMO
algunos de los factores más importantes controlados en el tronco del encéfalo son la presión arterial, la frecuencia cardíaca y la frecuencia respiratoria. En efecto, el corte transversal del tronco del encéfalo por encima de un nivel pontino medio permite mantener el control basal de la presión arterial sin cambios; pero impide su modulación por los centros nerviosos superiores, como el hipotálamo. Por el contrario, la sección inmediatamente por debajo del bulbo provoca su descenso hasta unos valores por debajo de la mitad de lo normal
Control de los centros autónomos del tronco del encéfalo por las regiones superiores Las señales procedentes del hipotálamo e incluso del cerebro tienen la capacidad de influir sobre la actividad de casi todos los centros de control autónomos situados en el tronco del encéfalo. Por ejemplo, la estimulación de las zonas adecuadas, sobre todo en el hipotálamo posterior, puede activar los centros de control cardiovascular bulbares con una potencia suficiente como para elevar la presión arterial hasta más del doble de lo normal. Análogamente, otros centros hipotalámicos controlan la temperatura corporal, aumentan o disminuyen la salivación y la actividad digestiva, y provocan el vaciamiento de la vejiga urinaria. Por tanto, hasta cierto punto, los centros autónomos del tronco del encéfalo actúan como estaciones de relevo para controlar las actividades iniciadas en niveles más altos del encéfalo, sobre todo en el hipotálamo.